前面我们介绍完了植物的营养器官——根茎叶,下面我们来看植物的繁殖reproduction过程。对于植物来说,繁殖方式多种多样,一般分为营养繁殖vegetative reproduction(植物的一定部位形成新的个体的方法,如块根、块茎和珠芽,以及具有克隆生长特性的植物的克隆系,农林上常会采用分株division、扦插cutting、压条layering、嫁接grafting来快速繁殖)、无性繁殖asexual reproduction(植株的一定部位产生具有繁殖能力的特化细胞或孢子spore,前者如有花植物的无配子生殖,后者如菌类植物、蕨类植物的孢子生殖)、有性生殖sexual reproduction(指配子结合为合子,再有合子发育为新的植物体,可以使后代有丰富的遗传多样性)。植物的花、果实、种子与植物的有性生殖有关,被称为生殖器官。下面我们就先来介绍花flower。

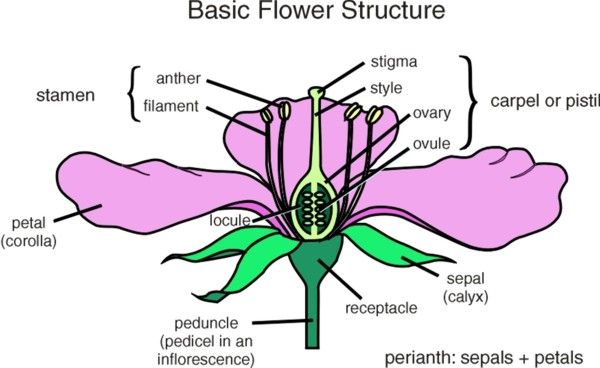
与叶类似,我们也可以将花分为完全花与不完全花,根据花萼和花冠(合称花被perianth)的有无,可以分为单被花、双被花、裸花。根据雌雄蕊的有无分为单性花、两性花、无性花(存在雌雄同株以及杂性同株)。花实际上是适应于生殖、极度缩短且不分枝的变态枝。花柄是枝条的一部分,花托通常是花柄顶端沿不同方向膨大的部分,花萼、花冠为不育的变态叶,雄蕊、雌蕊为可育的变态叶。
花柄pedicel:也称花梗,是连接花与茎的柄状结构,其基本构造与茎相似。既可以由茎向花疏松,又能支持着花。果实形成时,花柄发育为果柄。
花托receptacle:位于花柄的底端,是花器官其他各个组成成分着生的部位。其形态也随着植物种类而异,有些植物的花托为圆柱状、圆锥状、杯状、有的会扩大成为花盘desk,有的在雌蕊基部呈短柄状,传粉受精迅速伸长,将子房插入土中结实(比如花生),这种花托称之为雌蕊柄或子房柄。
花萼calyx:有一定数目的萼片sepal组成,常呈绿色,可以进行光合作用,为花芽提供营养;但也有一些植物的花萼呈现喇叭状,有利于植物传粉。花萼有离萼和合萼(其合生部位一般称为萼筒,未合生部位称为萼齿或者萼裂片)之分。有的植物在花萼之外还有一轮绿色的副萼epicaiyx。一般植物在开花后萼片就脱落了,但也有的在果实成熟后花萼依然存在,这种花萼被称为宿存萼,如茄。
花冠corolla:由若干花瓣petal组成,可以排列为一轮或多轮,由于有色体或液泡中的花色素所致,花瓣颜色比较鲜艳。有的花瓣基部可以分泌挥发油和蜜汁。花瓣彼此分离的花为离瓣花,花瓣彼此联合的为合瓣花,所以如同花萼一样也存在花冠筒和花冠裂片。
雄蕊stamen:由花药anther和花丝filament构成。花药是花丝顶端膨大为囊状的部分,内部有花粉囊,可以产生大量的花粉粒。花丝细长,基部着生在花托,或者紧贴在花冠基部
雌蕊carpel:多数植物的花只有一枚雌蕊。雌蕊由变态的叶即心皮pistil发育而来,由柱头stigma、花柱style、子房ovary三部分组成。柱头位于雌蕊的上部,是承受花粉粒的地方,多呈羽毛状,可以增加柱头接受花粉粒的表面积,多数植物的柱头可以分泌水分、脂类、酚类、激素和酶里帮助花粉粒附着和萌发。花柱位于柱头与子房之间,是花粉萌发后花粉进入子房的通道,与传粉的选择性有关。花柱提供花粉管生长的营养物质。子房是雌蕊基部膨大的部分,外为子房壁,内为一至多数子房壁。胚珠位于子房室内。受精后整个子房发育为果实,子房壁为果皮,胚珠发育为种子。由于组成雌蕊的心皮数目和心皮间的离、合情况的不同,雌蕊可以分为单雌蕊(一朵花只有一个心皮构成的雌蕊,其中离生单雌蕊是指一朵花中有多个彼此分离的单雌蕊)、复雌蕊(一朵花中只有一个由两个以上的心皮合生的雌蕊称为复雌蕊)
花在植物上的排列方式即为花序,花和花序均可以生于枝条的顶端或生于叶腋。植物体对成花所需条件起反应而必须达到的生理状态称为成花状态ripeness to state,而之前成为幼年期juvenility。植物经一定时期的营养生长后,在适宜的温度、光照和营养条件下,茎生长锥不再形成叶原基和腋芽原基,而发生花原基和花序原基。这一过程就是花芽分化flowerbud differentiation。花芽各部分的原基的分化顺序通常由外向内,最早出现的是萼片原基,以后依次向内产生花瓣原基、雄蕊原基、雌蕊原基,但是生长发育的快慢却有所不同。比如棉的花芽分化是副萼(苞片)原基迅速增大,后期发育成三个大型叶状副萼包于花外,内部出现五个萼片原基,萼片原基生长过程中,萼片原基基部彼此联合,上端保持分裂状态,形成5浅裂的花萼。花瓣原基与雄蕊原基为共同起源型。雄蕊管形成和生长,花芽的近中央部分出现3~5个心皮原基,之后心皮原基继续增大,相互愈合,分化出柱头、花柱和子房。而烟草首先是花序轴的分化和短暂生长,然后是花序分枝原基的分化,最后才开始花原基的分化。
而禾本科的花序是由小穗组成,小穗由小花组成,其分化为幼穗分化。小麦幼穗分化开始时,茎端生长堆迅速伸长,在两侧形成一系列环状苞叶基(单棱期)。接着从幼穗中下部开始,分别向上向下依次发育,在各苞叶原基的腋部分化出小穗原基(二棱期)。之后,小穗原基继续发育长大,苞叶原基逐渐消失。每一小穗中的分化顺序,是先在基部分化出两个颖片原基,之后再由下而上地进行小花原基的分化。小花的分化则依次形成一片外稃原基、一片內稃原基、两个浆片原基、三个雄蕊原基、一个雌蕊原基。小麦的雌蕊由两片心皮组成,雌蕊原基初发生时,心皮合生,成环状结构,包围着突起的单生胚珠,之后环状结构闭合,上部形成两个花柱,发展出羽毛状柱头。密议小穗上部小花的雌雄蕊时常退化不育。

而对于水稻,其幼穗分化与小麦有所不同。水稻为圆锥花序,当生长锥伸长,出现一系列的苞叶原基之后,在各苞叶原基腋部分化出的是一次枝梗原基,在一次枝梗上再分化出苞叶原基和二次枝梗原基。之后在一次枝梗顶端及二次枝梗上进行小穗原基的分化。其分化顺序为:先出现两枚颖片原基和两枚退化小花的外稃原基,再依次发育出小花的外稃、內稃、浆片、雄蕊和雌蕊的心皮原基。水稻小花中有六枚雌蕊,分为内外两轮,每轮各三枚。
总而言之,花芽分化与光周期、温度(如低温处理的春化作用)、基因(花器官分化的ABC模型)、外源激素(如赤霉素)。

花是植物体重要的生殖器官,而其中最关键的结构就是雌蕊和雄蕊。首先介绍一下雄蕊,我们前面提到,雄蕊是由花丝和花药组成。花丝结构简单,横切花丝,最外一层为表皮,中央有一个维管束,上连花药的药隔,下连花托,一般是在生长素(IAA)的诱导下以居间生长的方式迅速伸长。花丝有向花药转送营养物质、支持花药、促进开花,使花药暴露在空中,有利于花药的开裂和传粉。
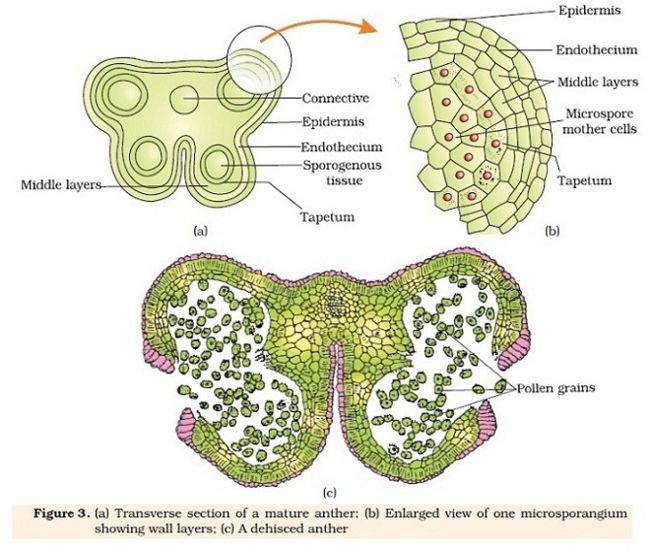
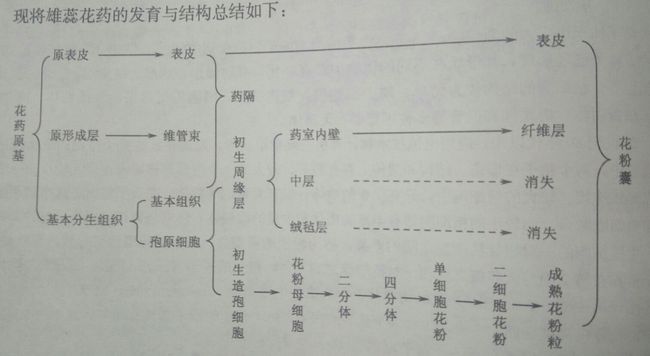
花药是雄蕊产生花粉的结构,由花粉囊(pollen sac)和药隔(connective)组成。多数植物的花药有4个花粉囊,花粉囊是产生花粉pollen grain的囊状结构,花隔是花药中部连接花粉囊的部分。花隔由通入花丝的维管束和周围的薄壁细胞组成。雄蕊原基经顶端生长和边缘生长基本完成幼嫩花药的发育,最初的一团分生组织,原表皮经垂周分裂形成将来花药的表皮,内方是基本分生组织,与药隔和花粉囊的发育,原基的近中央部分为原形成层,将来形成药隔维管束,与花丝维管束相连。幼小花药经过细胞分裂而迅速长大,由于四个角隅处的细胞分裂较快,横切面上的圆形变成了四棱形,之后原表皮层下面的第一层基本分生组织分化成为多列的孢原细胞archesporial cell,其细胞核较大,核大、质浓,分裂能力强。四个角隅处各有一个孢原细胞,之后进行平周分裂,形成内外两层,外层为初生周缘细层primary parietal layer,内层为初生造孢细胞primary sporogenous cell。花药中部的原形成层细胞逐渐分裂、分化形成维管束,并于其他基本分生组织发育而来的薄壁细胞一起形成药隔。初生周缘层细胞继续平周分裂和垂周分裂,自内向外逐渐形成药室内壁(endothecium)、中层(middle layer)和毡绒层(tapetum),与表皮一起构成花粉囊壁。药室内壁常为一层细胞,初期储藏大量淀粉和其他营养物质;毡绒层是花粉囊壁的最内层一层细胞,其细胞较大,初期为单核,中期可以形成多核细胞。毡绒层细胞质浓,细胞器丰富,对小孢子的发育和花粉粒的形成有重要的营养作用。初期造孢细胞经过分裂发育成花粉母细胞,在花药成熟过程中,花粉母细胞经减数分裂、发育成花粉粒。
成熟的花药由药隔和花粉囊两部分组成,花隔是连接花丝和花粉囊的重要结构,包括表皮、药隔基本组织和药隔维管束三个部分。表皮和药隔组织基本组织的淀粉等储藏物质被逐渐分散转移,药隔维管束进一步分化成熟,表现为导管壁增厚、腔明显。花粉囊(药室)由花粉囊壁(药室外壁与药室内壁)与花粉室(花粉囊腔和花粉粒)组成。花粉囊的外壁(即表皮)细胞呈扁长型,外壁可外凸,通常具有明显的角质层和气孔器。细胞内壁细胞细胞径向扩展,细胞壁内出现了以纤维素和少量木质素为主要物质的次生加厚,细胞内储藏物质逐渐消失,此时的药室内壁又称为纤维层fibrous layer。一些水生植物和闭花受精植物的药室内壁不发生带状加厚。一般会在两个相邻花粉囊的交接处不发生加厚,留下一条狭条状的薄壁细胞区域。开花后花药暴露,细胞失水,因纤维层外切向壁不增厚而产生较多的褶皱,机械组织使得临近的花粉囊交接处断开,形成裂口lacuna。中层细胞在花药成熟过程中,其细胞内的储藏物质被分解和转移,成为花粉粒发育成熟过程中的养分,其细胞壁被挤压变形逐渐解体并被毡绒层吸收和利用毡绒层在花粉母细胞开始减数分裂时,其细胞核分裂但不伴随细胞质的分裂,常形成具有双核和多核的细胞。前期从毡绒层生长旺盛。后期逐渐衰退。根据毡绒层发育后期的形态差别,可以将其分为腺质(分泌)绒毡层和变形(周原质团)毡绒层两种类型。可以提供花粉合成的原料、提供形成花粉外壁的孢粉素(孢粉素化学性质稳定,可以长久地保存在古代地层或泥炭层中)、合成分泌胼胝质酶、提供花粉外壁一种识别蛋白。花粉粒细胞会经过减数分裂形成四分体,四分体解离,发育成单细胞花粉,经1~2次细胞的有丝分裂逐步发育为成熟花粉粒。所以说成熟的花粉囊只剩下表皮、纤维层和成熟的花粉粒。
花粉母细胞在双子叶植物中为多边形,单子叶植物中多为近圆形,体积较大、细胞核大、质浓而且没有明显的液泡。花粉母细胞与毡绒层母细胞之间有胞间连丝存在,表明其结构和生理上保持联系,花粉母细胞逐渐积累胼胝质,形成胼胝质壁,并逐渐加厚至胞间连丝被阻断,原有的纤维壁消失。花粉母细胞发育到一定阶段开始减数分裂。其也分为连续型(多见于单子叶植物,第一次或者第二次分裂结束后随即形成细胞壁,四分体排列在一个平面上细胞壁的发育为离心式)和同时型(细胞壁发生在减数第二次分裂末为四面体型,细胞壁的发育方式为向心式)两种类型,之后多数植物胼胝质壁分解形成四个单独的小孢子,也有花粉块的存在。
花粉粒细胞初期细胞壁薄,细胞质浓,核位于细胞的中央,不断吸收毡绒层的分泌物,体积增大,细胞质中的小液泡合并为中央大液泡,细胞质和细胞核被挤在一侧,这一时期即为单核靠边期。之后细胞核在近壁处不均等分裂,其纺锤体多与花粉粒的壁垂直,细胞板弧形弯向生殖核的一侧。分裂结果形成一大一小两个细胞,大的为营养细胞(其功能与花粉发育中的营养以及花粉管的生长有关),小的为生殖细胞,两个子细胞之间的壁不含纤维素,主要由胼胝质组成。之后紧贴花粉壁的生殖细胞逐渐内移,细胞变圆,胼胝质壁消失,出现了细胞中有细胞的现象,之后生殖细胞逐渐变为纺锤形。而花粉壁在最初的单细胞花粉的胼胝质壁和质膜之间形成纤维素的初生外壁primexine沉积。质膜上形成圆柱形突起,穿过初生外壁辐射状排列于花粉粒的表面。外侧进一步积累孢粉素,形成基粒棒,基部扩展成为基足层foot layer,之后顶端扩展形成覆盖层tectum。覆盖层、基粒棒层共同构成花粉外壁的外层,基足层称为花粉外壁I,花粉质膜分泌纤维素形成花粉外壁II。花粉成熟后只有生殖细胞和营养细胞的花粉粒称之为二细胞花粉粒,还有一些植物如水稻、小麦、莎草还要进行一次有丝分裂,形成两个精细胞,成为三细胞花粉粒。所以成熟的花粉粒具有两层细胞壁,即内壁和外壁,内含2~3个细胞,即一个营养细胞和一个生殖细胞或者两个精细胞。
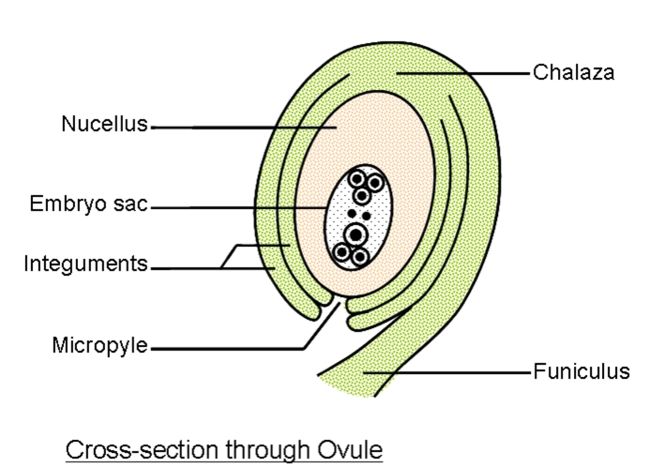
而雌蕊是由一至数个变态的叶卷合而成。组成雌蕊的变态叶称之为心皮carpel,每个心皮通常有三条维管束,其中相当于叶片中脉的维管束称为背束,两侧的维管束称为腹束。心皮在形成雌蕊时常常向内卷合,使得近轴的一面闭合起来,心皮边缘的连接处成为背缝线,雌蕊包括柱头、花柱和子房三个部分。其中柱头的表皮及乳突的角质膜外侧覆盖着一层亲水的蛋白质薄膜,可截获与识别花粉,有利于花粉萌发。根据开花时表面是否产生分泌物,我们将柱头分为干柱头(存在亲水性的蛋白质薄膜)和湿柱头(如可以黏住花粉并防止柱头失水的脂质,参与识别反应的蛋白质,防止侵害以及选择性促进花粉萌发的酚类,花粉粒萌发和花粉管生长的营养物质阿拉伯糖)。花柱可以分为空心型(有花柱道style canal存在,沟道表面有高度腺化的花粉道细胞,释放所产生的粘性物质)和实心型(花柱中央有富含细胞器代谢旺盛的引导组织transmitting tissue)。子房位于花托上,其结构由子房壁ovary wall(外层表皮上有许多的气孔和表皮毛,中层有多层薄壁细胞及维管组织系统,内层与外层相近但角质层分化不完全)、子房室locule(子房内的空腔,单雌蕊子房内只有一室,复雌蕊一至多)、胚珠ovule(生在胎座上的(腹缝线)上的卵形小体,是种子的前体)、胎座placenta组成。
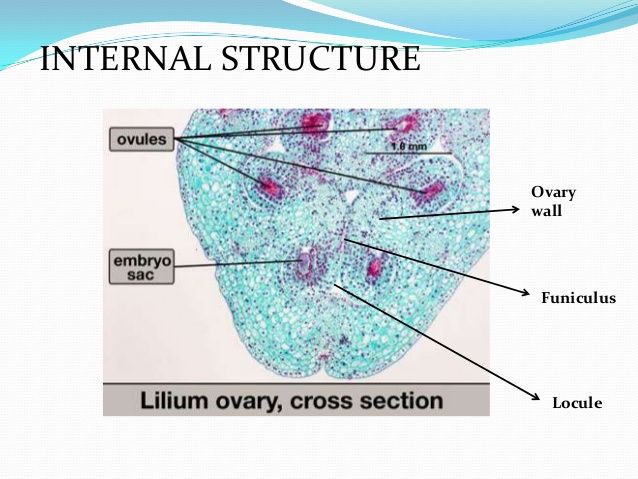
胎座表皮下层的细胞经平周分裂,突出成为胚珠原基,原基前端为珠心nucellus,是胚珠最重要的结构部分。胚珠中的胚囊是由珠心的细胞发育而成的。原基的基部发育成株柄funiculus。珠心基部的表皮层细胞可快速分裂,产生环状的突起,逐渐形成珠被micropyle。有的还有内珠被与外珠被之分,但内珠被发生较早,在珠心基部或者一侧,珠被、珠心和株柄汇合的一点称之为合点chalaza。心皮中的维管束分支从胎座经过株柄到达合点而进入胚珠内部,成为胚珠运输养料的途径。胚囊发育于珠心组织,开始形成时,在靠近珠孔的一端的表层下就发育出了一个与周围不同的孢原细胞archesporial cell,其细胞的体积较大,细胞质较浓,细胞器丰富,液泡化程度低,细胞核大而显著。之后平周分裂形成内外两个细胞,外侧的一个称为周缘细胞parietal cell,内侧的一个称为造孢细胞sporgenous 。发育为胚囊母细胞,周缘细胞继续垂周分裂和平周分裂,增加珠心的细胞层数,形成厚珠心。胚囊母细胞经过减数分裂,形成单细胞胚囊,可以分为:
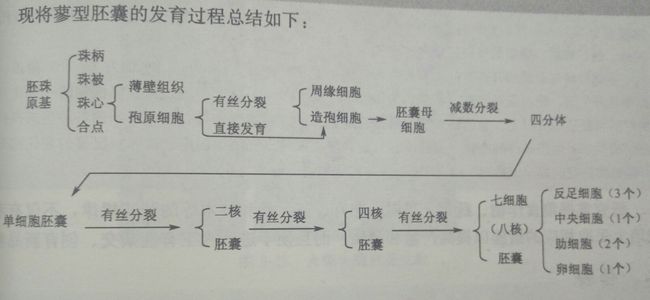
单孢型(蓼型):减数分裂形成四个单倍体的大孢子,之后沿着珠心呈线型或者T型分布,靠近珠孔的三个细胞相继程序性死亡。远离的那个发育为单细胞胚囊embryo sac。之后细胞核连续三次有丝分裂,第一次分裂形成两核移至胚囊两端,接着在进行两次分裂形成四核、八核胚囊。胚囊细胞逐渐发育长大,液泡化增强。在每端的4个核中,各有一个核向胚囊中部移动,互相靠拢,这两个细胞核称为极核polar nucleus。极核与周围的细胞质一起组成胚囊中最大的细胞,称为中央细胞central cell。在一些植物中,中央细胞中的两个极核常在传粉和受精前相互融合成一个二倍体的核,称为次生核secondary nucleus。近珠孔端的三个核,一个分化为卵细胞egg cell、两个分化为两个助细胞,它们常被合称为卵器egg apparatus,这两类细胞之间有胞间连丝协同配合。近合点的三个核会分化形成三个反足细胞antipodal cell。

双孢型(葱型):胚囊母细胞减数第一次分裂形成二分体,近珠孔端的一个细胞逐渐退化消失,近合点一端的一个完成减数第二次分裂,形成具有双核的单细胞胚囊,之后发育为成熟胚囊。
四孢型(贝母型):减数分裂只有核分裂没有细胞质的分裂。形成具有四个核的单细胞胚囊,再由具有4个核的单细胞胚囊发育成为成熟胚囊的方式。在四孢型胚囊发育的初期,近合点端的三个核,在随后的一次有丝分裂中,3个核的染色体先合并成两个体积较大的三倍体细胞,而珠孔端的一个核经过一次有丝分裂,形成8核胚囊。3个反足细胞和一个极核都是三倍体,而另一个极核、两个助细胞和卵细胞都是单倍体。
总而言之,成熟胚囊是被子植物的配子体,卵细胞称为雌配子。
一朵花中的雌蕊和雄蕊发育成熟,花萼和花冠开放,露出雄蕊和雌蕊,这个现象,就是开花anthesis。在开花过程中,雄蕊花丝迅速伸长并挺立,雌蕊柱头或分泌柱头液,或柱头裂片张开,或者羽毛状的柱头的毛状物突起,有利于接受花粉。而花药中的花粉散出,借助外力传到雌蕊柱头上的过程称为授粉或者传粉pollination。分为自花授粉(在进化生物学上有繁殖保障效应,最典型的自花传粉为闭花授粉和闭花受精,可以使花粉粒在外界不良的水湿条件下能够正常传粉受精和种群繁衍)和异花授粉。植物在长期进化过程中在生理和形态上其生理和形态构造上形成了许多适应于异化传粉的特性:单性花(unisevual flower)(雌雄同花—>雌雄异化同株—>雌雄异株)、自花不孕(花粉粒落到自花的柱头上不能萌发,或者花粉管生长缓慢)、雌雄异熟(一朵花上的雌雄蕊成熟时间不一致)、雌雄异长(同种或者同种群的不同个体间产生的两种或者三种两性花)、雌雄异位(花内[同种个体只产生一种两性花,花中雌雄蕊高度不同]或者花间)、曲卷性花柱(花柱通过反向弯曲,回避自身的花药),传粉的方式也有风媒和虫媒两种。
受精是指雌雄配子相互融合的过程。首先花粉与柱头之间存在某种识别反应,对亲和性好的花粉柱头会提供水分、营养物质等物质刺激花粉萌发。花粉内壁分泌角质酶,溶解柱头表皮细胞的角质层。如果不亲和柱头乳突细胞会产生胼胝质将萌发孔阻塞,组织萌发。亲和的花粉产生水合反应,从柱头的分泌物中吸收水分和营养,内壁从萌发孔处向外突出,形成花粉管,向花柱和子房生长若为二细胞花粉,生殖细胞和营养细胞随之进入花粉管先端,营养细胞向前,生殖细胞分裂一次,形成两个精子,三细胞花粉中的营养细胞和两个精子都进入花粉管之中。花粉管的生长从突破柱头开始可以沿着花柱道的粘性分泌物生长,实心花柱中常沿着引导组织或者细胞壁中生长。花粉管通过花柱,到达子房后,一般沿着子房的内壁或者胎座继续生长,直达胚珠。通常花粉管是从珠孔进入胚珠为珠孔受精,而有些植物的花粉管通过合点区或者珠被进入胚珠,称为合点受精。
多数被子植物的花粉管到达胚珠之前或进入胚珠后,胚囊中的两个助细胞中的一个常先退化,花粉管通过胚囊的壁,精细胞、一个营养核、细胞质等花粉管内含物一起喷泄而出。形成特定的细胞质流,将精细胞带到卵和中央细胞所在位置,其中一个精子与卵细胞结合形成二倍体的合子,将来发育成胚,另一个精子与极核结合形成三倍体的初生胚乳核,将来发育为胚乳。这个现象为双受精。之后花粉管裂口被胼胝质堵塞,防止胚囊中其他物质倒流入花粉管。如果有多条花粉管进入胚囊,这样胚囊里就有了两对以上的精子,这就是多精子现象。多于的精子会被胚及胚乳吸收,起到营养作用。有时的细胞会使得助细胞或者反足细胞受精,出现了多胚现象,有时也会发生两个以上的精细胞入卵,发生多精入卵现象。
参考:
https://en.wikipedia.org/wiki/Flower
https://en.wikipedia.org/wiki/Pedicel_(botany)
https://en.wikipedia.org/wiki/Calyx
https://en.wikipedia.org/wiki/Stamen、
https://en.wikipedia.org/wiki/Carpel