自从ETR1基因被成功克隆后,关于乙烯受体方面的研究就开始层出不穷。人们逐渐发现,乙烯受体并非“一枝独秀”,而是由多个“兄弟”组成了一个受体家族。我们可以把ETR1比作乙烯受体家族创业的开拓者,它完成了最原始的积累,并带领着其他成员一路开疆拓土,创建了显赫的乙烯受体家族。下面就让我们一起扒一扒乙烯受体家族的“发家史”吧。
在上一讲中,我们曾经提到过,Elliot M. Meyerowitz实验室1993年发表的克隆ETR1的Science文章中发现了一个非常有意思的现象:当用ETR1基因和拟南芥基因组DNA杂交时,可筛选到多条片段,说明ETR1可能存在一个基因家族。那么这些基因是什么,它们和乙烯信号转导到底有什么关系呢?在接下来的几年里,Elliot M. Meyerowitz实验室对这一科学问题进行了深入细致的研究。
1995年,Elliot M. Meyerowitz实验室在Science上发表文章Ethylene insensitivity conferred by Arabidopsis ERS gene。他们以ETR1的cDNA为探针,对拟南芥基因组DNA文库进行筛选,筛选到一个同源基因,命名为ERS(ethylene response sensor)。ERS定位在2号染色体上,编码613个氨基酸,和ETR1蛋白具有67%的一致性(identity),但是比ETR1短了125个氨基酸。ERS的N端(1-321)和ETR1的相似性为75%,在蛋白质数据库中没发现已知结构域注释;C端含有一个预测的组氨酸激酶结构域,和ETR1具有58%的相似性,但是少了一个receiver domain。值得注意的是,在对乙烯反应突变体的正向遗传学筛选中,一共筛选到了4个ETR1等位突变(etr1-1至etr1-4),这4个突变位点分布在3个疏水结构域中,而在ERS中,这4个位点都是保守的。
虽然ERS和ETR1具有一定的相似性,但是它在乙烯信号转导中发挥什么作用呢?在etr1-4突变体中,ETR1蛋白第62的Ile被Phe替换,导致显性的乙烯不敏感。既然这个位点在ERS中也是保守的,那么ERS中发生相同的突变后,会发生什么呢?科研人员利用定点突变的方法,将相同的突变引入ERS中,并转化拟南芥。和etr1-4相同,转化ers的拟南芥黄化苗也表现出显性的乙烯不敏感表型,并且在其他生长时期同样也都对乙烯不敏感。以上结果说明,ERS可能具有和ETR1类似的乙烯受体功能。

在对拟南芥乙烯反应突变体筛选的过程中,Elliot M. Meyerowitz课题组还筛选到了另外一个显性的乙烯不敏感突变体etr2-1。1998年,相关研究成果以ETR2 is an ETR1-like gene involved in ethylene signaling in Arabidopsis为题发表在The Plant Cell上。通过图位克隆,鉴定到了位于3号染色体上的ETR2基因。etr2-1编码区发生了一个C到T的突变,导致第66位的氨基酸由Leu突变为Pro,而这一位点处于乙烯受体家族非常保守的区域。将etr2-1基因转化到野生型拟南芥中,导致转基因植株出现显性的乙烯不敏感表型,说明etr2-1突变体的表型确实是由ETR2基因突变所致。
对蛋白序列分析发现,ETR2编码773个氨基酸,和之前鉴定的乙烯受体ETR1以及ERS具有较高的相似性。蛋白结构方面,ETR2和ETR1具有相同的结构域:一个N端结构域,一个可能的组氨酸激酶结构域以及一个接收结构域(receiver domain),而ERS的接收结构域缺失。但是,就蛋白序列的相似性而言,ETR1和ERS的相似性更高(79%),而ETR2与ETR1和ERS的相似性相对较低(65%和63%)。

通过前面的研究,已经鉴定到了3个可能的乙烯受体,但是基于以下证据,Elliot M. Meyerowitz等仍认为乙烯受体家族可能还有新的成员:
1. 之前筛选到了一个ein4突变体,它具有和乙烯受体突变体相似的乙烯不敏感表型,但是EIN4的基因定位和功能方面的研究还未开展;
2.由于许多乙烯反应突变体只筛选到了一个allele,而且并没有筛选到ERS突变(ERS通过交叉杂交技术筛选得到),因此乙烯反应突变体的遗传学筛选应该还没饱和。
鉴于以上两点,Elliot M. Meyerowitz课题组通过反向遗传学的方法,利用交叉杂交(cross-hybridization)技术,对新的乙烯受体家族成员进行了筛选。1998年,相关成果以EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis为题,发表在The Plant Cell上。
文章中以ETR2的cDNA为探针,对拟南芥的基因组进行筛选,筛选到了两个ETR2的同系物(homolog),其中一个定位到3号染色体上。有意思的是,之前通过遗传学方法筛选到的ein4突变体,图位克隆定位到的候选区间和该同系物的候选区间有重叠。并且ein4突变体的表型和之前鉴定到的3个乙烯受体突变体相似,都是显性的乙烯不敏感,因此科研人员猜测这个ETR2同系物很可能就是EIN4。另外,通过正向遗传学筛选,又筛选到了两个ein4的等位突变ein4-2和ein4-3。进一步研究发现,该ETR2同系物确实就是EIN4。EIN4编码766个氨基酸,和ETR2具有较高的相似性(74%)。
另外一个ETR2同系物定位在1号染色体上,被命名为ERS2,它的氨基酸序列和ETR2以及EIN4的相似性较高(73%和68%)。和EIN4不同,之前并没有通过遗传学方法筛选到ERS2的等位突变体,为了验证基因的功能,通过定点突变的方法将ERS2蛋白第67位的Pro突变为Leu(etr2-1具有相同突变),94位的Ile突变为Phe(etr1-4,ein4-1和ers1-1具有相同突变),并转化至拟南芥中。转基因植株在T1代就表现出乙烯不敏感的表型,并且在自交的T2代中,乙烯不敏感表型和转基因是共分离的,说明ERS2突变可导致显性的乙烯不敏感。
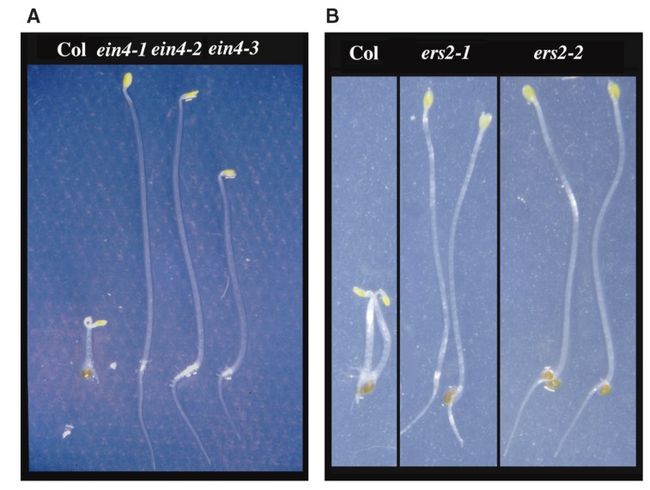
通过以上研究,一共找到了5个可能的乙烯受体,对这5个基因的表达模式进行分析后发现,它们的时空表达存在一定的差异性。并且,ETR1和EIN4的表达对乙烯处理没有响应,而乙烯处理后ERS1、ERS2以及ETR2的表达量增加。以上结果说明,这5个疑似的乙烯受体在乙烯信号转导方面可能存在功能差异。
虽然已经找到了5个候选乙烯受体,但是有一点需要引起我们的注意:之前筛选出的乙烯受体突变体都是显性突变,而显性突变有可能是由于loss of function引起,也有可能是由于gain of function引起,所以并不能确定这些基因在乙烯信号转导中到底起到正向作用,还是反向作用。因此,筛选以上基因的loss of function突变体才能最终确定基因的功能。
还有一个问题,为什么通过遗传学方法没有筛选到受体的loss of function突变体呢?可能有以下三个原因:1.以上受体基因对植物的生长发育是必须的,任一基因发生loss of function突变致死;2.乙烯受体家族可能存在功能冗余,某一受体突变后,并没有明显表型;3.以上基因可能并不参与乙烯感知,它们loss of function后并不改变植物的乙烯反应。
因此,通过筛选以上受体基因显性突变的基因内抑制子的方法,也许能够筛选出loss of function突变体。所谓基因内抑制子,在此处是指在发生显性突变的同一基因内部又发生了另一突变,抑制或部分恢复原突变体乙烯不敏感的表型。使用基因内抑制子筛选的方法,Elliot M. Meyerowitz课题组成功鉴定到了乙烯受体loss of function的突变体,相关结果以Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana为题于1998年在Cell上发表。
通过EMS诱变的方法,成功筛选到了ETR1、ETR2和EIN4的loss of function突变体,这些突变体都是发生了提前终止或者剪切错误,为隐性突变,可恢复原突变体乙烯不敏感的表型。另外,鉴于在之前的诱变中没有筛选到ERS2的突变体,因此采用筛选T-DNA插入突变体库的方法,筛选到了ERS2 loss of function突变体。

对这些loss of function突变体的乙烯反应表型研究后发现,单突并没有明显的乙烯反应缺陷表型,单突杂交后得到的双突表型也不明显,直到三突和四突才表现出明显的组成型乙烯反应的表型。说明乙烯受体家族存在严重的功能冗余,这也解释了为什么之前没有筛选到乙烯受体loss of function突变体。由于乙烯受体loss of function突变体三突和四突表现出组成型的乙烯反应,这说明和大多数受体的正调控机制不同,乙烯受体在乙烯信号转导通路中起到负调控的作用。也就是说,在没有乙烯时,受体处于激活的状态;当有乙烯时,受体的活性反而被抑制。另外,由于乙烯受体loss of function突变体表型和显性突变体表型相反,因此显性突变应该属于gain of function。

2005年,Bleecker实验室在The Plant Journal上发表了一篇题为Ethylene-binding activity, gene expression levels, and receptor system output for ethylene receptor family members from Arabidopsis and tomato的文章,发现拟南芥的五个候选乙烯受体和乙烯分子都具有高亲和的结合活性。
至此,就集齐了乙烯受体家族基因作为乙烯受体的三大证据,确立了乙烯受体家族的地位:
1. 这些基因的突变体都可导致植株的各个组织和器官在所有生理时期都表现乙烯反应缺陷的表型;
2. 在所有已知的乙烯信号组分中,这些候选受体基因都位于遗传学上游(具体内容后续会逐步介绍);
3. 具有特异的乙烯结合活性。
通过以上解读我们发现,乙烯受体开创性的研究大部分都是Meyerowitz实验室完成的。其实,这种现象在科学研究中是非常常见的。一个实验室专注于某一领域,进行深入系统的研究,建立非常完善的体系。科学研究其实就像一个连环案,一个案件破解了,往往又引出下一个案子的线索。而科学家们就像福尔摩斯,用他们那一双慧眼在错综复杂的事件中发现关键线索,破解一个又一个扑朔迷离的案件背后的真相,数十年如一日,乐此不疲。
【参考文献】
Chang, C., Kwok, S. F., Bleecker, A. B., & Meyerowitz, E. M. (1993). Arabidopsis ethylene-response gene ETR1: similarity of product to two-component regulators.Science,262(5133), 539-544.
Hua, J., Chang, C., Sun, Q., & Meyerowitz, E. M. (1995). Ethylene insensitivity conferred by Arabidopsis ERS gene. Science, 269(5231), 1712-1714.
Sakai, H., Hua, J., Chen, Q. G., Chang, C., Medrano, L. J., Bleecker, A. B., & Meyerowitz, E. M. (1998). ETR2 is an ETR1-like gene involved in ethylene signaling in Arabidopsis. Proceedings of the National Academy of Sciences, 95(10), 5812-5817.
Hua, J., Sakai, H., Nourizadeh, S., Chen, Q. G., Bleecker, A. B., Ecker, J. R., & Meyerowitz, E. M. (1998). EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis.The Plant Cell,10(8), 1321-1332.
Hua, J., & Meyerowitz, E. M. (1998). Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell, 94(2), 261-271.