原文: Microbiota and Obesity: Where Are We Now?
Biology (Basel). 2020 Dec; 9(12): 415. Published online 2020 Nov 25.
doi: 10.3390/biology9120415

摘要:
-
简单总结
国际科学文献中报道的新数据表明,人类肠道菌群的特定改变是肥胖症和与肥胖有关的代谢性疾病的特征。 肥胖受多种因素制约,微生物群无疑是重要的因素。 通过对实验研究数据的分析,我们可以假设微生物群落组成的变化可能是人类代谢变化的原因,而不仅仅是其结果。 需要进行广泛的样品临床试验,研究饮食诱导的肠道菌群对宿主代谢的调节作用,以了解在分子水平上的相互作用,从而观察到代谢与菌群变化之间的相关性。
-
摘要:
遗传和环境因素是肥胖和其他代谢疾病的根本原因,因此很难找到合适且有效的药物治疗。 然而,毫无疑问,肠道菌群以及口腔中存在的细菌是这些病理发展的关键因素,但其机理尚未得到充分描述。 ·当然,对微生物群结构的更详细的了解(组成,种间和种间关系,代谢功能)可能对抵抗肥胖的发作有很大帮助。 识别关键细菌种类将使我们能够创建“健康”细菌的数据库,从而可以根据代谢和临床需求来操纵细菌群落。 ·在临床护理中将肠道菌群定位为肥胖症和与健康相关的并发症(甚至只是减肥)的治疗已成为现实可能性。
- 在本专题综述中,我们概述了微生物群在宿主能量稳态和与肥胖相关的代谢性疾病中的作用,因此探讨了新的和现有策略(营养/饮食调节和粪便微生物群移植的影响)的代谢性疾病的治疗治疗潜力。
- 关键词:微生物群,与肥胖有关的代谢性疾病,营养不良,益生菌和益生元,粪便微生物群移植,临床生物化学和临床分子生物学,免疫系统,临床微生物学,营养和体育锻炼对健康的影响
1.简介
- 近年来,我们目睹了全世界肥胖人群的流行趋势:根据最新数据,据估计,全世界超重或肥胖人群超过20亿[1],到2030年,美国成年成年人中有42% 人口有望肥胖[2]。 成人肥胖症与生活质量受损和许多健康问题相关,包括代谢综合征,糖尿病,癌症,呼吸系统和心血管疾病,以及易患代谢综合征 。
- 微生物群是人类新陈代谢的关键因素,最终会影响我们的健康或反过来影响我们的健康,并有利于各种病理的发生和发展。 人们普遍认为,包括口腔微生物在内的微生物群与肥胖症的发展有关[5]。 口腔微生物群与肥胖之间的相关性已在许多科学论文中得到报道,其始于不同的细菌组成[6],可导致味觉改变[7],有利于食用所谓的舒适食品,通常热量高。
我们所食用的每种食物中未消化的成分成为微生物群的食物来源,这反过来又使我们获得了其他代谢产物,这些代谢产物不仅对能量收集很重要,而且对于它们在调节和信号传导途径中的作用也很重要。 越来越明显的是,微生物组在各种病理(慢性和传染性)的出现和发展中具有基本作用。 因此,将微生物群落作为识别和治疗各种病理情况的临床策略的潜在目标是很合乎逻辑的。 尽管对微生物群和肥胖的关系进行了大量的研究,但是在分子水平上仍然没有明确的解释[8]。 - 最近已经在小鼠中证明了有希望的分子联系,但是人类肥胖症和相关代谢功能障碍的多种因素(包括遗传学/表观遗传学和生活方式)使得难以证明肠道营养不良的独立作用。 生物科学完全能够表征微生物群落的各个方面(例如,组成,功能,产生的代谢产物,物种之间的关系),但无法清楚地确定这些方面对代谢可能发展的贡献改变和任何病理,因此可能用于治疗目的。
2.肠道菌群
- 在科学文献中已经确定,肥胖个体和健康个体的微生物群在组成和功能上都存在实质性差异[9]。 毫无疑问,饮食习惯(例如,西方饮食或限制性饮食)可能有助于改变这些参数,增加新陈代谢的压力,进而导致肥胖[10]。 肥胖症中的微生物群失调症的特征是微生物群落多样性显着降低,例如,以Firmicutes与Bacteroidetes的比例增加为特征[11]。 有趣的是,如果受试者体重减轻,可以改变改变的比例[11,12,13]。
这些组成差异反映在微生物群代谢功能的重大变化中:在肥胖个体中,短链脂肪酸(SCFA)产生者(Actinobacteria)和致病性Proteobacteria大量增加,表明这种细菌可能在 肥胖者的代谢变化[14]。 关于微生物组的组成,宏基因组学分析表明,肠道微生物基因库中有40%是全世界所有个体之间共享的,因此支持了核心微生物组的存在(表1) - 尽管由于使用的算法不同而产生的结果不同,但这一概念还是值得商,的,这是微生物群指纹,微生物随时间推移的稳定性的概念,而不管所有变化的可能性(即年龄,生活方式,性别,昼夜变化) (久坐,身体活动等)—可以保护包括能量代谢在内的几种重要功能途径[16,17]。
表格1
-
在人体肠道菌群中占主导地位主要的细菌门和属:
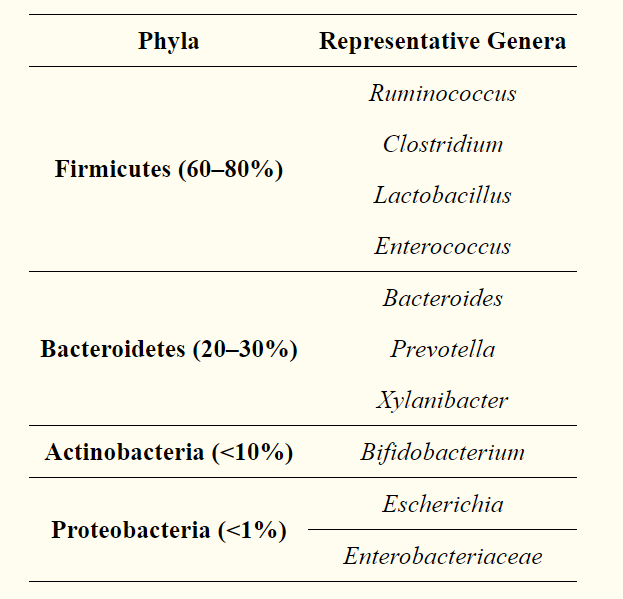 image.png
image.png 诸如人类微生物组项目和美国肠道项目之类的项目在肠道微生物组分析中可能会非常有帮助,有助于我们阐明微生物在健康和疾病状态中的作用和关系。 除了肥胖以外,在多种病理中都发现肠道菌群改变,包括肠道肠综合征(IBS),2型糖尿病(T2D) 和心血管疾病[24],帕金森[25]。 真正的问题是给构图的变化赋予绝对和因果值; 最近的研究发现了不同的变化,因此排除了肥胖发展的明确分类标志 [26]。
将肠道菌群与肥胖联系起来的分子机制仍然难以捉摸,但包括从食物中获取能量,肠道屏障通透性和免疫系统功能以及产生影响能量代谢和信号传导途径的特定代谢产物,从而对人体代谢产生明显影响。
肠道菌群的营养调节会影响总能量摄入,养分吸收,运输和储存,这反映在一般宿主代谢的下游,最终有益于健康或促进体重增加。 例如,那些经常食用高纤维和多酚饮食的人,其微生物群的多样性更大,可以被定义为实质上更健康,而西方饮食的消费者则发现其丰富度较低 **。药物和抗生素治疗是影响肠道菌群组成的另一个重要因素。 微生物群的组成和功能受到化学疗法的深远影响,受到损害,有利于机会性感染的出现[28]。 二甲双胍是一种用于糖尿病治疗的著名药物,能够直接作用于微生物群。 然而,应该强调的是,简单观察作为肥胖病因的不同细菌门之间的关系变化具有明显的科学局限性。 这可能是肥胖的结果,而不是肥胖的原因,而且,它没有充分考虑物种或菌株内部可能发生的角色变化,即使水平基因转移也是如此。 高物种多样性与基因含量/多样性甚至健康状况并不相同。 元基因组学研究清楚地突出了微生物群的代谢潜力比简单的生物分类观察中存在的物种复杂得多。 因此,尝试全面了解肥胖与肠道菌群失调相关的分子机制将更为合适。
3.代谢功能及其与宿主代谢的相互作用
-
微生物与我们的新陈代谢之间的相互作用比最初想像的要大:在饮食生活方式和遗传背景的相互影响下,细菌群落可以直接或间接地影响能量收集,免疫功能,激素分泌,肠屏障通透性甚至情绪和神经系统。 微生物群在维持肠道屏障功能中起着重要作用,肠道屏障功能对我们的健康至关重要。 该屏障可防止潜在的病原细菌和抗原进入,并最终竞争营养和粘附,从而避免了炎症状态和免疫系统细胞的募集[30]。 含微生物群核苷酸结合的低聚结构域蛋白(NOD)1和2帮助淋巴组织预防炎症并促进上皮细胞再生[31]。
关于包含N末端吡喃结构域(NLRP)NLRP1,NLRP3和NLRC4炎性小体的nod样受体,已证明这些炎性小体的上游信号相对较好,并且既可以是微生物来源的,也可以是宿主来源的( 图1) 。
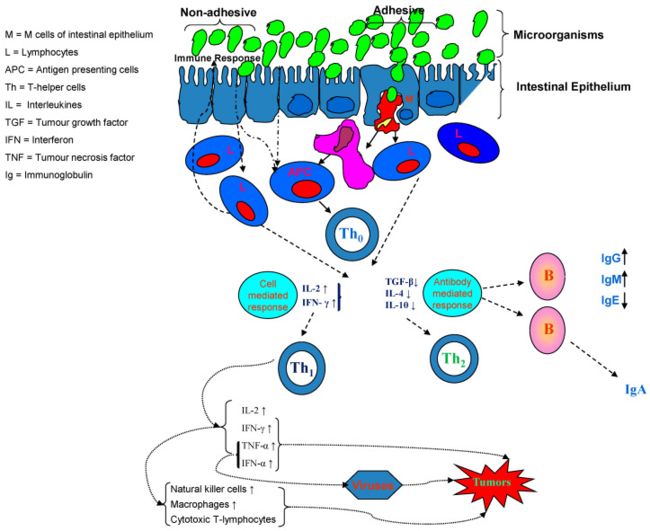 image.png
image.png
尤其是,NLRP3炎性小体的独特之处在于它对各种刺激(包括细菌和非微生物刺激物)产生响应,例如细菌成孔毒素,颗粒物,氧化的线粒体DNA和细胞外ATP [33]。
最近,NLR家族成员NLRP6被认为能够通过自噬途径调节杯状细胞功能,并能够响应细菌信号而促进Muc2分泌,这两者对于维持完整的粘液层(提供有效的 防止细菌性炎症的屏障[34]。 微生物及其代谢产物是通过氨基酸,抗菌化合物和酚类衍生物的生产在结构和功能上塑造肠道屏障的关键因素[33]。胆汁酸的代谢在很大程度上受到肠道微生物群组成的影响。 胆汁盐水解酶(BSHs)是将不同种类的细菌(拟杆菌属,双歧杆菌,梭状芽孢杆菌,乳酸杆菌)产生的用于将初级胆汁酸转化为次级胆汁酸的共轭作用的关键酶。 其他胆汁酸反应(7α-脱羟基,酯化和氧化/表位化)也需要肠道菌群,最终控制脱氧胆酸的水平,进而抑制其受体,主要是法尼类X受体(FXR)和武田G蛋白偶联受体。 FXR和TGR5是调节人类整体代谢的基础:FXR调节肝脏中脂肪酸的摄取,合成和氧化; TGR5(也与FXR合作)促进胰高血糖素样肽1(GLP-1)的分泌,从而导致胰岛素分泌增加和胰高血糖素合成减少,调节骨骼肌的能量消耗和胰岛素敏感性,并具有抗炎作用 。
值得注意的是,丁酸(其在营养不良症中的水平降低)可以上调GLP-1R的表达。
补充丁酸钠可以预防NAFLD患者从单纯性脂肪变性发展为脂肪性肝炎[37]。值得注意的是,饮食健康对微生物群体的影响也比肥胖个体对微生物群落的影响要大得多。 饮食产品(例如海鲜或鱼类)过多会增加肠内细菌三甲胺(TMA)的产生,可能与心血管疾病有关[38]。
通过筛选与心血管疾病(CVD)相关的代谢产物来鉴定TMA及其产物三甲胺N-氧化物(TMAO),并且TMA需要肠道细菌来形成[39]。 在结肠中形成并吸收后,TMA进入门脉循环,将血液引导至肝脏,在肝脏中,其被含黄素的单加氧酶3(FMO3)氧化为TMAO [37]。 口服抗生素可阻止通常在胆碱或肉碱饮食攻击后发生的TMAO的增加,表明TMAO的产生需要微生物细菌[38]。
饮食是改变小鼠微生物结构的主要原因,而不是遗传背景 。 研究表明,遵循西方饮食的人的微生物群具有非常特殊的特征,其结构发生了深刻的变化,即使在物种或基因水平上也没有明显的差异[35]。
此外,每种饮食方式似乎都有其独特的特征,与其他饮食不同。 Sonnenburg及其同事[36]的实验表明,不适合微生物群落的饮食会破坏微生物群落,导致故意和费力地在整个生命中产生物种和菌株的消失,有时甚至一代代。 不幸的是,仅靠饮食的改变(改善)并不能弥补微生物的损失:只有通过使用不同的技术,尤其是粪便微生物群移植(FMT),才有可能恢复小鼠的原始微生物群丰富性。 使用这些研究作为克服细菌损失和多样性差的模型,有效的策略可以理解益生菌的补充或FMT。
4.肥胖和其他代谢疾病
菌群失调与多种疾病有关:肥胖,代谢综合症,糖尿病,肠肠疾病,肝病如肝硬化和NAFLD,以及癌症; 他们都显示出与肠道菌群变化的相关性[37]。
肥胖与微生物群之间有什么联系? 在2000年代初期,来自Bäckhed及其同事的无菌(GF)小鼠的开创性实验有助于建立肥胖与微生物群的关系[38]。
用常规饲养的肥胖小鼠进行微生物群移植后,GF小鼠表现出新陈代谢改变,即使在热量限制的情况下,其胰岛素抵抗能力和脂肪量也持续增加[39]。 数据清楚地表明了微生物群在能量收集中的作用,即使在热量限制的情况下,尤其是微生物群移植能够改变受体小鼠的几种代谢特征[39]。 微生物群失衡与超重和肥胖之间的关系可能是由于许多因素引起的。 多年来,已经提出了各种机制的假设:当然,微生物群代谢产物似乎起着重要作用。 SCFA约占我们新陈代谢的能量资源的10%[40]。 与非肥胖个体相比,在超重和肥胖个体中,SCFA含量要高得多(这可能是热量过多的结果)[38]。 这是一个极其重要的方面,不仅是从纯粹的精力充沛的角度来看,而且还因为它们作为信号分子的潜在作用,能够在宿主的新陈代谢中产生功能性变化。 例如,SCFA与葡萄糖稳态有关。此外,不应低估SCFA的抗炎作用-通过组蛋白脱乙酰基酶(HDAC)抑制,Toll样受体以及通过刺激T调节细胞的分化[40]。 因此,某些新陈代谢的后果可以通过观察到的SCFA产量增加来解释[36]。 SCFA可能参与葡萄糖稳态(提高葡萄糖敏感性)和通过游离脂肪酸受体(FFAR2 / FFAR3)进行脂质代谢,从而导致AMP激活的蛋白激酶活化。
代谢后果刺激肝糖异生和脂肪生成途径的激活[40],同时抑制肌肉中的脂肪酸氧化[41]。
肠道通透性的增加会增加LPS的摄取并导致全身性炎症[42]。 在小鼠身上进行的实验表明,存在阿克曼粘菌,粘液层和肠道通透性降低之间存在直接联系,从而防止了炎症过程[42]。 在严重胰岛素抵抗的db / db小鼠中,每天用哈氏杆菌L2-7进行为期四周的治疗不会产生不良影响,并且可能通过改变丁酸酯的形成,胆汁酸和代谢而对代谢产生有益的影响。在另一项研究中,无菌C57BL / 6J肠杆菌诱导的肥胖小鼠与从志愿者肠道分离出的阴沟肠杆菌B29株单联显示血清内毒素负荷增加和炎性疾病加重[44]。 这种人类来源的内毒素生产者在生糖小鼠中的肥胖诱导能力表明,它可能是导致其人类宿主肥胖发展的原因[44]。
肠细菌可以差异地调节脂质代谢。 确实,脱硫弧菌分泌的产物上调CD36的表达,而梭菌产生的产物则下调CD36的表达。 因此,丧失调节CD36表达功能的生物体可能导致脂质吸收不当,脂质会随时间累积,导致肥胖和代谢综合症[45]。
研究表明,髓样蛋白表达的NLRP12通过减弱脂肪组织中的TNF,IL-6,NF-kB,MAPK,M1巨噬细胞极化和炎症小体活化来抑制高脂饮食诱导的肥胖和2型糖尿病。
NLRP12的这种保护功能取决于微生物群,并且与鞭毛藻科及其代谢产物有关,从而减轻了肥胖[46]。 除了在能量代谢中发挥作用外,SCFA在肠屏障水平上对防御病原体也很重要。 在小鼠实验中,由于梭状芽胞杆菌的损失,丁酸含量低会改变结肠细胞的氧合作用状态,进而促进机会性细菌的感染[43]。 来自肥胖个体的脂肪细胞的炎性信号可能损害肠屏障的完整性和功能,促进病原体及其促炎分子的转移[44]。 因此,微生物群改变可以促进与肥胖有关的慢性低度炎症状态[45]。过氧化物酶体增殖物激活受体γ(PPARγ)可能通过触发FIAF(空腹诱导的脂肪因子)激活而参与了另一种可能的机制,该激活是一种对脂蛋白脂肪酶(LPL)具有抑制作用的肠上皮细胞蛋白,脂蛋白脂肪酶是脂质代谢的关键因素。 微生物群也参与FIAF表达的调节:微生物群失调可以抑制FIAF表达,从而导致脂肪组织中LPL活性和脂质积累增加[47,48]。 肥胖,代谢疾病和较低的微生物多样性之间的联系已经在一些研究中阐明。 与非洲或南美的农村地区人群相比,分析美国城市人群的粪便微生物菌群得到了证实,美国城市人群的纤维消耗量低且肥胖率高[49]。
使用宏基因组学分析,TwinsUK队列研究(世界上最大的成年双胞胎注册和临床上最详细的研究)发现,多样性较低与腹部肥胖有关[50]。 总而言之,即使个体中肠道菌群的组成不均匀,肥胖个体中微生物多样性的降低似乎也是一个一致的特征。
当然,这些变化会影响微生物群代谢产物的产生,对能量代谢,肠屏障功能和炎症问题产生显着影响。 SCFA的产生和代谢的改变似乎是微生物群失调引起的后果的一个反复出现的主题。然而,其他细菌代谢产物在能量稳态,免疫调节系统以及防御和炎症发展中也很重要。
-
遗传 ,年龄,体育活动和许多其他因素对肠道菌群的功能有重大影响,因此很难建立微生物群在肥胖症发展中作用的因果关系[51]。在小鼠中,体育锻炼会促进肠道菌群的多样性[52]:低强度运动会限制肠内短暂的大便时间,从而降低结肠癌,憩室病和炎性肠病的风险。 在人类中,肠道微生物的多样性与心肺健康有关[53]。 衰老的进程与人类微生物组多样性的降低有关,这是老年人中通常观察到的脆弱的可能原因[54]。 识别这些变化背后的分子修饰可以极大地帮助人们更有效地对抗衰老的后果,甚至即使使用适当的益生菌也是如此。 食物中的动植物组织会在许多步骤中被各种细菌降解。
短链脂肪酸,甲烷和乙酸盐是由与食物分解有关的细菌产生的。 这些产品然后打开或关闭宿主基因和其他邻近微生物的基因。 粘膜层覆盖绒毛,也为粘膜微生物的长期居住提供了一个家。当粘膜微生物在粘膜层中繁殖时,它们在宿主的整个生命周期中都停留在该处,并且很少离开。 相反,随着变化的发生(例如,食物供应,食物量,病原体),管腔微生物种群的数量和类型可以迅速变化(图2)。
 image.png
image.png
图2
- 微生物群落相互作用的简化版本:肠壁在底部,绒毛和血液供应将氧气带入肠壁。 图的顶部显示了肠的中段/腔,其氧供应低,但来自内腔中的硫酸盐还原细菌的硫化氢供应更高。
5.食物和微生物群:相互影响
- 肠与调节食欲的中枢神经系统之间存在着密切的关系。 营养状况,代谢能水平,神经和肠内肽的分泌(后者也取决于肠道中食物的存在)和激素信号都是调节卡路里摄入的因素[55]。 已经显示出一些细菌菌株能够影响肠道激素的合成,例如YY,GLP-1,瘦素和生长素释放肽,调节大脑的食欲和饱腹感[56,57]。 肠脑轴(GBA)是人体中枢神经系统(CNS)与肠道神经系统(ENS)之间的双向链接。 它涉及大脑中具有外周肠功能的认知和情感中心之间的直接和间接途径。 GBA涉及内分泌下丘脑-垂体-肾上腺轴(HPA),免疫(细胞因子和趋化因子)与植物神经系统(ANS)之间的复杂串扰[57]。 GBA主要结合了ANS的交感神经和副交感神经,分别驱动肠道和大脑之间的传入和传出神经信号。 同时,HPA轴可协调针对压力的适应性反应,包括激活大脑边缘系统的记忆和情绪中枢[58]。 GBA的神经,免疫,内分泌介质可使大脑影响肠道功能(免疫细胞,上皮细胞,肠神经元和平滑肌细胞)。
*此外,胃肠道(GI)的细胞也受到肠道菌群的影响,最近的证据表明,微生物组在GBA结构中起着重要作用[59,60]。 微生物来源的代谢产物,例如短链脂肪酸,可以与肠内分泌细胞上的受体结合,从而改变肠激素向体循环的释放[58,59]。
富含非消化性碳水化合物的饮食可以提高动物和人类的SCFA水平和肠道激素水平[58]。
微生物群代谢产物的作用非常广泛,尚未完全了解。 确实存在由神经系统介导的肠,肠神经系统和大脑之间的双向串扰,这对调节我们的新陈代谢至关重要。
值得注意的是,微生物群能够合成代谢产物,例如5-羟色胺和γ-氨基丁酸(GABA)[60],它们在中枢神经系统中活跃,它们参与控制食欲,从而间接地影响体重。 醋酸盐(一种由肠道细菌产生的SCFA)能够通过抑制食欲刺激而在下丘脑中发挥作用。
Byrne及其同事证明,在啮齿动物中静脉内补充乙酸盐可影响食欲中枢的食欲信号,从而抑制食欲刺激[61]。有趣的是,肥胖症中的微生物群失调可以提高乙酸盐水平,促进葡萄糖刺激的胰岛素分泌,增加生长素释放肽的分泌,进一步增加肥胖症[64]。 在精神病学中,疾病和肥胖症经常联系在一起:毕竟,焦虑或沮丧的人在食物中寻求安慰是一种普遍的经验,经常选择热量摄入高的食物[65]。 因此,微生物群在某些神经系统显示中起着重要作用,可能还因为它充当了大脑与饮食之间的纽带:从具有心理障碍的小鼠身上移植微生物群的实验也使受体小鼠沮丧 。
-
乳酸杆菌和双歧杆菌的许多种都产生GABA,GABA是大脑中主要的抑制性神经递质。
**此外,念珠菌,大肠埃希菌和肠球菌可产生神经递质5-羟色胺,而某些芽孢杆菌已显示产生多巴胺。 细菌还产生SCFA,例如丁酸,丙酸和乙酸,它们能够刺激交感神经系统,粘膜5-羟色胺释放,从而影响大脑的记忆和学习过程 **。
微生物群与宿主代谢之间的共生关系是众所周知的。 米尔斯及其同事的研究表明,人乳中存在的低聚糖(HMO)的第一受益者是细菌,而不是新生儿。 事实证明,婴儿双歧杆菌能够使用HMO作为独家食物来源生长[69]。 因此,人类母乳和婴儿双歧杆菌之间的相互作用类似于在母亲与新生儿的关系中招募新的参与者-细菌本身。 不幸的是,将这些多种相互作用转化为对我们的健康有益的饮食适应症是极其复杂和困难的。
在特定的细菌菌株和我们的代谢改变之间建立直接联系相对容易。 但是要证明与益生菌之间的这种特殊联系无疑是具有挑战性的(或者与益生菌之间的联系更为复杂)。
6.饮食干预和微生物群调节
饮食和微生物群绝对与人类健康和疾病息息相关。 现在我们可以肯定地知道我们的生活方式,尤其是我们的饮食习惯如何能够在我们的肠道菌群中产生深刻的变化,从而改变其组成,生态和功能[55]。 越来越多的证据表明,富含多酚的饮食(主要存在于植物来源的食品和饮料中)可以对人体产生抗氧化和抗肥胖作用[27,70]。 健康均衡的饮食,其中还包括每天正确摄取卡路里,在当今的西方世界,这是一个难以追求的目标,它通过不同的分子机制对我们的健康产生积极影响,其中一些分子机制直接涉及微生物群
我们的微生物群及其显着特征(组成,功能,生态)受许多因素的影响,有些因素无法改变(出生方式,基因型,年龄,衰老),但许多因素却可以由我们的行为改变:体育活动,久坐的生活方式, 周围环境,温度等[71]。 当然,我们的营养在塑造微生物群中起着根本性的作用:据估计,它会影响多达60%的细菌菌群组成[72]:我们的饮食对微生物群落产生深远影响,并根据需要对其进行塑造(表2) 。
表2

- 很明显,热量过高或脂肪,碳水化合物和/或蛋白质含量过高,例如所谓的西方饮食,如果随时间不断延长,将决定微生物菌群的决定性结构变化。 甚至我们不吃的东西(纤维,微量营养素,矿物质)也可以显着改变肠道细菌。 没有膳食纤维的饮食(因此没有纤维中存在的易消化的碳水化合物的进入)通常会导致细菌菌群多样化的减少[34,82]。
·这些数据已通过对小鼠进行高脂肪和高糖饮食的实验得到了证实,例如经典的西方饮食方案使小鼠变胖并改变了微生物群,改变了Firmicutes(高代谢简单碳水化合物的能力)和Bacteroidetes之间的比例 。饮食习惯的重要性通过最小化微生物群变化来确认,无论是否将小鼠带回限制体重增加的饮食,都强调了微生物群代谢功能与营养之间的紧密联系。 - 食物与微生物的相互作用产生了一个基本问题:是否可以通过饮食预防疾病?
·暂时的答案尚不清楚; 也许应该,但是一定要为每个人量身定制 。
6.1 热量限制
- 热量限制似乎是我们可以使用的重要工具:最近有研究表明,仅40%CR的一个月就可以改善胰岛素敏感性,降低体重(BW)和增加白色脂肪的质量,而没有任何明显的不利影响[84]。 在这些条件下分析肠道菌群显示组成逐渐变化,这可能是由于胰岛素敏感性提高和肥胖症减少所支持。 有趣的是,在相同的饮食和热量摄取条件下,移植了CR-菌群的小鼠与对照组相比,体重减轻了。 终生CR可提高与预期寿命呈正相关的细菌菌株的百分比[85],而在系统水平上,LPS结合蛋白(LBP)的血清水平明显降低。
·CR可能通过负面调节其生物合成而影响血液中LPS的水平。 微生物群的改变显着促进了CR期间许多代谢变化:在CR期间小鼠中发现的较低SCFA浓度[86]可能有助于改善CR期间观察到的葡萄糖能量代谢。 除了较低的SCFA产生(尤其是丁酸和丙酸),还可以防止刺激脂肪细胞增殖[87]和减弱饱腹感信号[88]。
6.2 益生菌和益生元
- 实际上,科学文献中充斥着许多关于益生元或益生菌的施用报告,以出于健康目的来修饰,营养和培养肠道菌群。 益生菌是对宿主有益的活微生物,而益生元是能够积极影响肠道菌群生长和活性,最终修饰宿主代谢的各种途径(包括能量,调节和免疫系统)的生物活性物质。 这里真正的问题是这些影响是否矛盾。
益生元的使用与微生物群变化之间是否存在直接的因果关系还是仅仅是其他变化的后果[89],仍然存在争议。 益生元被定义为肠道微生物群直接利用的一组营养素。 它们促进细菌菌株的生长,发育和功能,从而有助于微生物群落本身和人体的健康 (表3)。
表3

- 它们具有改变宿主代谢的多种作用:口服益生菌可以改善特应性皮炎的症状[95],通过TLRs受体调节免疫调节细胞,主要作用于碳水化合物代谢的调节[96]。 然而,益生元对微生物群的功效的有效性有许多方面需要考虑,从施用益生元的类型开始,以食物中存在的剂量开始,最后以人类代谢调控的多样性结束。 肠道菌群是我们健康的关键组成部分-食用含有益生菌的食物的数量已大大增加,也许希望能够治疗或改善一系列疾病。
一些益生菌菌株在克罗恩病的治疗中可能很有前途[97]。 - 但是,所引用的文章均未找到可推荐使用的大量证据[98]。 将益生菌科学转化为益生菌食品需要面临一些科学挑战:根据最新研究,个人微生物群和肠道粘膜定居的差异会大大改变益生菌摄入的预期结果[99]。 为了支持益生菌介导的微生物组变化为宿主带来健康益处,需要更多的证据:大多数关于益生菌对肠道菌群变化影响的研究已在动物模型中进行; 对人类的临床试验是有限的,有时是矛盾的[100]。
- 益生菌已被用于治疗与微生物组破坏有关的疾病,但是,尽管鼓励了体外实验[81],但它们在体内的功效却有限[101]。 此外,可以低估施用益生菌的适得其反的风险:它甚至可能减慢人类志愿者体内所需肠道菌群的恢复 。
7.针对微生物群的治疗策略
提出关于肠道菌群改变和对人类有益的健康后果的有用策略是非常具有挑战性的。
真正的问题是,考虑到我们自己的独特性,如何评估什么是健康的微生物组,以及应该对哪种细菌菌株进行鉴定,尽管显然在科和目层面上已经确定了与肠道菌群相一致的微生物群 。此外,关于肠道菌群的最重要问题应该集中在哪些物种真正具有代谢活性,从而调节宿主的新陈代谢,而不用担心各个物种的相对丰度。 它应该是非常有用的诊断工具,可将微生物组特征用作疾病存在的生物标记。
众所周知,所有代谢性疾病都涉及某种类型的微生物群失调,两者都是因果关系,但这绝非易事。 关于微生物群可能对我们的健康有益的另一项研究是使用(或促进产生)由细菌群落成员产生的用于治疗目的的小分子抗炎药。 它们调节宿主信号传导途径的能力和宿主生理学是一个令人着迷的话题,但仍有待阐明。 肠道细菌代谢产物可通过调节特定的宿主信号传导途径来提供生理稳态。 为了清楚地阐明理解肠道菌群与人类健康和微生物群相关疾病之间关系的关键概念,需要使用不同的动物模型进行进一步的研究。
肿瘤病理的发生和发展似乎与我们的微生物群有直接关系,尽管其分子机制尚待确定。众所周知,某些细菌来源的代谢物[105]能够触发促癌或抗癌途径:例如,某些SCFA(例如丁酸)可能在某些细胞系(腺癌细胞)中具有这种作用,改变细胞周期并最终导致细胞凋亡 。 在乳腺癌中,肠道微生物群衍生的代谢产物在调节癌细胞功能以及建立和维持肿瘤微环境中很重要[108]。 显然,饮食是这些代谢产物的主要来源。
例如,高脂肪和高蛋白饮食是现代西方饮食的特征[109,110],这是发生癌症的危险因素之一。 因此,假设富含某些食物会促进产生SCFA的细菌的新陈代谢的饮食,可能会导致某些对象的肿瘤性疾病的发作,这似乎是正确的。好消息是,在塑造肠道菌群的组成,结构和功能时,生活方式,饮食和体育锻炼明显超过了我们所继承的任何遗传学。 未来的目标是利用一种精密的药物:针对每个患者的病情个性化微生物替代疗法的可能性,从而使干预措施尽可能有效。 Elinav及其同事的最新研究[99]表明,益生菌在人类患者中没有相同(或可再现)的作用。 经过四周的补充剂整合后,所收集的微生物群落样本显示,只有部分人的消化道中发现了有益细菌。
在另一些细菌中,细菌仅存在于粪便样本中,而不存在于消化道中[113]。在过去的几十年中,减肥手术一直是选择在极端情况下体重增加的选择。 修改胃肠道系统的解剖结构,该干预措施可调节养分的转运并影响肠道菌群状态[114]。
长期以来,FMT被视为一种非常有前途的技术。 实际上,它是艰难梭菌感染治疗的特权技术[115]。 可能有可能为FMT干预选择最合适的供者-受者匹配。 最近,FMT还被建议作为癌症治疗以及与癌症治疗相关的并发症的可能工具。 初步报告显示,FMT可以刺激癌症的免疫治疗效果,从而显着改善患者的临床预后[116]。 我们对微生物群及其与代谢的关系的了解的进步,将使我们能够根据个人及其个性特征,提出不断优化的疗法,从而选择最合适的细菌菌株来改善患者的肠道生态系统, 对新陈代谢有更大好处。
如今, 宏基因组学,元转录组学和元蛋白质组学以及生物信息学工具等新技术已帮助我们识别细菌群落,突出了健康微生物群与肥胖微生物群的差异。代谢组学是对包括代谢产物的非蛋白质小分子的研究,不仅可以帮助我们识别存在的物种,而且可以阐明物种之间的代谢关系以及它们与宿主代谢的相互作用 。 16 s rRNA基因构成了微生物群落分子研究的基本基础(图3A,B) 。
图3

- 为了更有效地预防和治疗特定的病状,正在开发替代技术,称为生物疗法,以用于粪便供体。 例如,粪便的人工合成(从选定的细菌菌株开始,通过发酵在体外产生的活细菌代谢产物)实际上正在临床试验中,似乎非常有前途[117]。
8.讨论
- 肥胖和与肥胖相关的代谢性疾病具有人类肠道菌群的特定改变。 肥胖受多种因素制约,微生物群无疑是重要的因素。 通过对实验研究数据的分析,我们可以假设微生物群落组成的变化可能是人类代谢变化的原因,而不仅仅是其结果。 如果这些前提是正确的,那么细菌菌群可能是预防或治疗与肥胖有关的疾病的可能干预措施的理想目标 。
- 代谢组学和代谢组学研究可以真正改善我们的知识,促进微生物群操作作为临床疗法。 正如一些饮食干预研究所表明的那样,肠道菌群的调节可以改善人体内的代谢参数,但是尚未确定肠道菌群在此类实验中的因果作用。
- 如前所述,有许多变量可以极大地改变个性化营养策略的结果:寻找饮食干预与代谢变化之间的真正关联仍然是科学家要实现的目标。 需要进行临床试验,以研究饮食诱导的肠道菌群调节对宿主代谢的作用; 这些分析将帮助我们了解分子水平上的相互作用,以观察到我们的代谢与微生物群变化之间的相关性,阐明肠道微生物群的饮食调节是否真的可以诱导宿主代谢中的代谢变化,从而改善人体健康 。
- 但是,应该强调的是,“与健康有关的”微生物群可以各种形式存在,例如结构,组成和功能。 因此,尽管有共同的特征,但我们每个人都会有适合自己特定需要和特征的个人微生物组。
去:
9.结论
- 超重和肥胖也因其对卫生系统的影响而成为全世界非常严重的问题; 因此,制定有助于减少损害的策略至关重要。 微生物群肯定与这种情况密切相关:细菌菌群的分析是评估或自身代谢的重要指标,但最重要的是,它可以是改善个人健康的理想目标。 考虑到使用益生元的益处和几乎完全没有风险,使用益生元和益生菌肯定必须成为抗肥胖全球战略的一部分。
- 新出现的技术或现有程序的改进(例如FMT)对于帮助减肥和/或改善肥胖者的临床状况可能至关重要:未来的科学进步可能使微生物群恢复治疗成为一种可能。 一般而言,与肥胖和代谢性营养不良有关的疾病患者的治疗中通常选择。