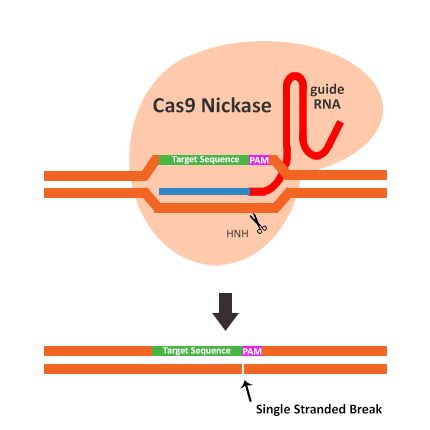
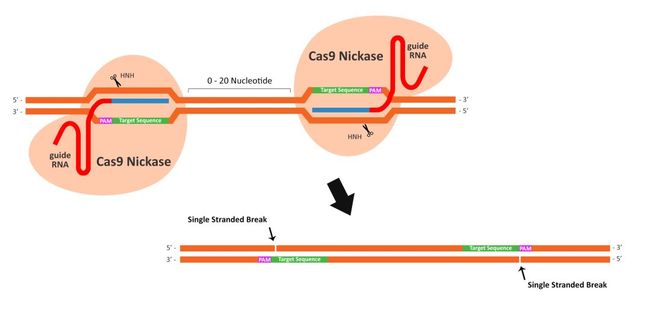
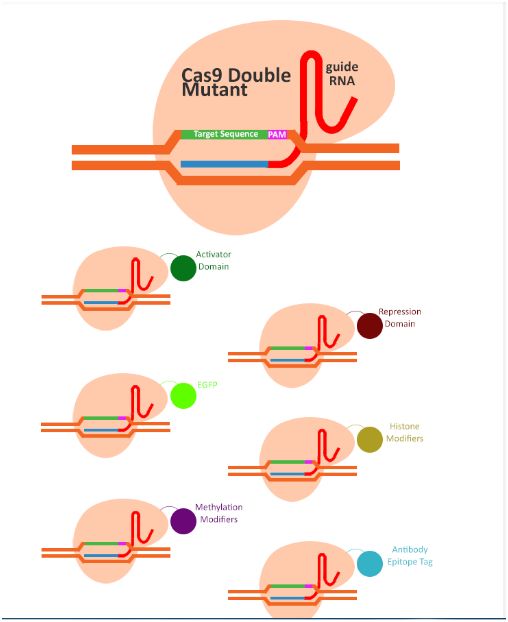
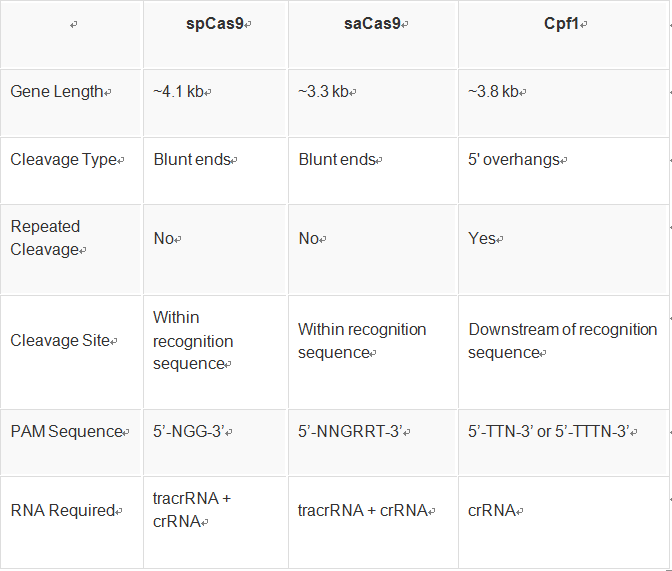
- 数据库的后悔药:Undo Log揭秘
你一身傲骨怎能输
游戏行业领域知识专栏撤销日志(UndoLog)
文章摘要撤销日志(UndoLog)是数据库的“后悔药”机制,用于保证数据操作的原子性和一致性。其核心原理是修改数据前先记录原始状态到UndoLog,若事务失败则进行回滚恢复。典型应用包括:1)事务回滚(如转账异常时还原数据);2)并发控制(通过快照读提供多版本视图)。主流数据库如MySQLInnoDB和Oracle均采用该技术,其流程可概括为“先备份后修改,出错即还原”。简言之,UndoLog通过
- 高效沟通05-FFC赞美法则
古城码农
工作记实录高效沟通沟通技巧
效沟通专栏–组织运转的命脉与个人成功的基石目录1.概念1.1底层逻辑1.2优势1.3适用场景1.4技巧2.示例1.概念 FFC是由三个英文单词首字母组成的结构化赞美模型,旨在让赞美真实可信、打动人心,避免空洞敷衍。其核心是通过三个层次构建逻辑闭环:要素含义作用关键要点F(Feeling)感受表达主观情感反应用情绪词传递真诚(如“感动”“佩服”)例:“你的方案让我非常惊喜!”F(Fact)事实提供
- react组件内点击事件的this的4种指向方法
程序员--韩同学
reactreact.jsjavascript前端
目录方法一.通过bind改变点击事件内的this指向外部组件内this方法二.通过在构造函数constructor内使用bind对函数内的this重定向方法三.通过箭头函数在事件内改变this指向方法四.通过使用箭头函数来指向外部组件内this(使用较多)扩展:1.react组件内点击事件传参2.Event事件,获取元素本身总结方法一.通过bind改变点击事件内的this指向外部组件内thison
- 产品经理高效工作指南,核心能力全拆解!
AI大模型-大飞
产品经理人工智能大数据智能体程序员大模型大模型学习
在互联网行业的激烈竞争中,产品经理作为产品的“操盘手”,其工作流程的科学性与专业性直接决定着产品的成败。想要高效推进产品从0到1、实现从1到N的迭代,必须吃透日常工作的每个环节。今天,我们就用思维导图为你拆解产品经理9大核心工作流程,从需求到迭代全链路解析,助力你成为更专业的PM!一、需求分析与市场调研:产品的“方向盘”需求分析是产品工作的起点,决定着产品是否贴合市场。市场调研:定期研究行业动态(
- 这是gpt o1给出的物联网工程专业的大学规划,有人看看这个合理吗?
王倚山
gpt物联网学习开发语言
下面是一份更为详细、覆盖全年(包括寒暑假)的四阶段学习规划,旨在帮助你在大学剩余时间里持续学习、循序渐进地掌握物联网(IoT)核心技能,打造深厚的技术壁垒。每个阶段都有明确的学习目标与自学内容细节,并在寒暑假安排了“强化期”任务,让你全年不停歇,不断提升。总体思路稳扎稳打:从嵌入式基础到RTOS、传感器驱动、通信协议,再到边缘计算、云平台、工业协议、安全攻防,层层深入。项目驱动:每个阶段至少完成1
- 养老机构运营实训室建设要点:构建实战化运营管理实训体系
凯禾瑞华_实训室建设
实训室建设大数据物联网智慧健康养老服务与管理虚拟仿真教学人工智能智慧养老
养老机构运营实训室作为养老服务人才培养的重要载体,其建设质量直接影响专业人才的实践能力与行业适配度。围绕实战化运营管理实训体系的构建目标,需从多维度精准把握建设要点,打造契合行业需求的实训环境。点击获取实训室建设方案一、明确建设目标与定位(一)贴合行业需求养老机构运营实训室建设要点的核心,在于精准对接养老行业发展趋势与实际需求。随着老龄化社会加速,养老服务精细化、智慧化需求激增,实训室应锚定培养具
- 交换机端口及VLAN转发原理
hao_wujing
网络
交换机端口及VLAN转发原理是数据通信网络的核心基础。理解它们的工作原理对于设计、管理和排错网络至关重要。下面我将详细解释:##一、交换机端口基础交换机端口是物理连接点,用于连接终端设备(如PC、服务器、IP电话)或其他网络设备(如另一台交换机、路由器、防火墙)。端口的主要职责是**在数据链路层(OSI第2层)转发以太网帧**。1.**关键概念:*****MAC地址表:**交换机内部维护一张表,记
- 卵生兄弟网络工作原理和架构
hao_wujing
安全
大家读完觉得有帮助记得关注和点赞!!!以下从网络安全角度解析**卵生兄弟网络**的深度技术架构与工作原理。该概念通常指**具有冗余唤醒和协同攻击能力的模块化恶意网络**(如APT组织所用),或**防御端的双生活动目录/蜜罐系统**。我们将从攻击与防御两个维度展开:---###一、攻击视角:恶意「卵生兄弟网络」架构(APT高级威胁)####**核心设计目标**-**隐蔽持久化**:主模块被清除时,备
- 基于虚拟化技术的网闸安全交换:物理隔离时代的智能数据流通引擎
109702008
#linux系统安全安全人工智能网络
摘要:在等保2.0和零信任架构背景下,传统网闸正从“物理断网”向“智能交换”演进。本文将深入解析如何通过硬件虚拟化+策略容器化在网闸内部实现安全数据交换,并提供工业级落地方案。一、痛点:隔离与效率的终极矛盾当企业面临以下场景时,传统网闸力不从心:生产网与办公网需实时同步数据库公有云与私有云间敏感文件传输多租户环境下跨安全域业务协同核心矛盾:物理隔离阻断攻击链的同时,也阻断了业务流!二、技术破局:虚
- Python知识点:如何使用memory_profiler进行内存分析
开篇,先说一个好消息,截止到2025年1月1日前,翻到文末找到我,赠送定制版的开题报告和任务书,先到先得!过期不候!如何使用memory_profiler进行Python代码内存分析在开发高性能的Python应用程序时,理解和优化内存使用是至关重要的。memory_profiler是一个强大的工具,它可以帮助你监控Python代码的内存使用情况。本文将介绍如何使用memory_profiler来分
- JavaScript基础语法之运算符和控制流
AA-代码批发V哥
JavaScriptjavascript
JavaScript基础语法之运算符和控制流一、运算符1.1算术运算符:数值计算的基石1.1.1字符串拼接陷阱1.2比较运算符:条件判断的起点1.2.1严格比较(`===`)vs松散比较(`==`)1.2.2其他比较运算符1.3逻辑运算符:复杂条件的组合1.3.1短路逻辑(重要特性)1.3.2实战:表单验证1.4赋值运算符:数据存储的桥梁1.4.1基础赋值(`=`)1.4.2解构赋值(ES6新增)
- Linux基础第5天-:Vim编译器的常用指令
今天也好累
linuxvim运维笔记学习编辑器服务器
光标移动(了解)行间移动gg键:移动光标到第一行(命令模式下)G键:移动光标到最后一行(命令模式下):n:移动到第n行,:6(移动到第6行)(末行模式下)列间移动$键:移动光标到当前行的行尾(最后一列),一般可以使用Shift+$(命令模式下)0键:移动光标到当前行的行首(第一列)(命令模式下)方向键↑↓上下实现行间移动,←→左右实现列间移动删除(重点)列删除x键:删除当前光标所在出的一个字符(命
- react中的this指向问题
匀升ovo
reactreact.jsjavascript前端
react中this指向问题对于class来说,在class中定义的方法实际上绑定在类的原型中(prototype),我需要通过class的实例调用这个方法才能拿到实例的this例如下方代码classPersonextendsReact.Component{constructor(props){super(props);this.state={name:props.name}}render(){r
- React this指向问题
state//1.创建组件classWeatherextendsReact.Component{//构造器调用几次?————1次constructor(props){console.log('constructor');super(props)//初始化状态this.state={isHot:false,wind:'微风'}//解决changeWeather中this指向问题//在生成的对象上加上
- 大语言模型应用指南:ReAct 框架
AI大模型应用实战
javapythonjavascriptkotlingolang架构人工智能
大语言模型应用指南:ReAct框架关键词:大语言模型,ReAct框架,自然语言处理(NLP),模型融合,多模态学习,深度学习,深度学习框架1.背景介绍1.1问题由来近年来,深度学习技术在自然语言处理(NLP)领域取得了显著进展。尤其是大语言模型(LargeLanguageModels,LLMs),如BERT、GPT系列等,通过在大规模无标签数据上进行预训练,获得了强大的语言理解和生成能力。然而,预
- 【Python】memory_profiler
宅男很神经
python开发语言
1.1引用计数与垃圾回收:Python的“贴身管家”与“清洁工”Python,特别是其标准实现CPython,其内存管理的核心是建立在一个优雅而高效的组合机制之上的:以引用计数为主,分代垃圾回收为辅。1.引用计数(ReferenceCounting):主要的内存管家这是CPython内存管理的基石。其原理极其简单:CPython中的每一个对象(一个整数、一个列表、一个自定义类的实例),其内部都维护
- 2019年架构师系列教程:高并发Netty实战打造百万连接架构
不教书的塞涅卡
本文还有配套的精品资源,点击获取简介:本课程面向高级IT专业人士,旨在教授如何利用Netty框架设计和实现能够处理高并发连接的服务器架构。Netty是一个高性能、异步事件驱动的Java网络应用程序框架。课程将提升学员在系统架构设计和性能优化方面的技能,应对高并发场景挑战,特别是在金融、游戏、物联网等领域。1.Netty框架基础概念介绍Netty是一个高性能的网络应用框架,专为快速开发可维护的高性能
- Python 数据分析实践:车辆行驶数据处理心得
lzzy-lt-0415
python数据分析开发语言
在数据驱动决策的大趋势下,Python凭借其丰富的数据分析库,成为处理各类数据的得力工具。近期我围绕车辆行驶数据展开分析,过程中收获诸多实战经验,在此分享用Python进行数据处理与分析的心得,也结合代码讲讲实际运用思路。一、数据导入与初步探索:开启分析第一步importpandasaspd#导入数据df=pd.read_excel(r'../../数据层/数据集合/车辆行驶记录表单2.xlsx'
- JavaScript基础语法之变量声明和数据类型
AA-代码批发V哥
JavaScriptjavascript
JavaScript基础语法之变量声明和数据类型一、变量声明1.1变量声明的本质1.2三种声明方式对比(var/let/const)1.2.1var:函数作用域的“老派选手”1.2.2let:块级作用域的“新生代”1.2.3const:常量声明的“守护者”二、数据类型2.1原始数据类型(PrimitiveTypes)2.1.1字符串(String)2.1.2数值(Number)2.1.3布尔(Bo
- SOC的工作原理和架构
hao_wujing
安全
大家读完觉得有帮助记得关注和点赞!!!SOC(安全运营中心)是企业安全防御体系的**神经中枢**,通过整合技术、流程和人员,实现**7×24小时安全监控、威胁检测、事件响应及持续改进**。其核心目标是**缩短攻击驻留时间(MTTD/MTTR)**,将碎片化的安全数据转化为可行动的防御策略。以下是SOC的工作原理与架构深度解析:---###**一、SOC的核心工作原理**####**1.安全数据全域
- Android之Application的onTerminate能监听应用退出吗?
zhangphil
AndroidAndroid
Android之Application的onTerminate能监听应用退出吗?一些Android开发者在不经意间发现Android的Application中有一个公开的回调方法:onTerminate()继而想当然的认为该方法即是Android的整个App应用退出后的回调,因为Terminate的词面意思就是结束,终结嘛。那么onTerminate在Android的App全部结束后回调,真的是这
- 【PyTorch】教程:torch.nn.GELU
老周有AI~算法定制
PyTorchpytorch深度学习python
torch.nn.GELU原型CLASStorch.nn.GELU(approximate='none')参数approximate(str,optional)–gelu近似算法用none或者tanh,默认为none;定义高斯误差线性单元函数GELU(x)=x∗ϕ(x)\text{GELU}(x)=x*\phi(x)GELU(x)=x∗ϕ(x)其中ϕ(x)\phi(x)ϕ(x)为高斯分布的累积分布
- Android PNG/JPG图ARGB_8888/RGB_565解码形成Bitmap在物理内存占用大小的简单计算
AndroidPNG/JPG图ARGB_8888/RGB_565解码形成Bitmap在物理内存占用大小的简单计算Android的Bitmap是一个用于表示图像数据的核心类,代表一张图片在内存中的存储,Bitmap存储了图像的像素信息数据。Bitmap把图像理解为像素点组成的二维矩阵,每个像素点存储对应位置的一系列ARGB值(透明度+红绿蓝通道)。Bitmap在内存中占用大小的关键计算公式:内存
- 数据结构之栈实验
lannnn_
学习记录数据结构c语言栈
栈实验实验目的实验环境实验要求实验内容源代码运行结果实验目的掌握栈这种数据结构特性及其主要存储结构,并能在现实生活中灵活运用。实验环境CodeBlocks实验要求1.熟悉c语言的语法知识;2.掌握栈的顺序存储结构—顺序栈的定义、构造、获得栈顶元素、入栈、出栈等基本操作;实验内容完成栈的定义、构造、获得栈顶元素、进栈、出栈等函数的编写。要求在主函数中实现对以上操作的调用,编写一个算法判断给定的字符向
- 守护线程及定时器
小白白成长记
多线程程序人生经验分享java
守护线程(后台线程)在java语言中线程分为ldalei用户线程和守护线程(后台线程)其中守护线程代表有垃圾回收线程守护线程的特点一般守护线程是一个死循环,所有的用户线程结束,守护线程就结束(main方法也是一个用户线程)*守护线程的用处假设每天00:00时候系统数据自动备份这个时候就需要设置定时器,并且可以将定时器设置为守护线程定时器定时器的作用是间隔特定的时间,执行特定的程序在Java的类库中
- 基于锁的获取与释放方式即计划于所得获取与释放方式进行分类——显式锁和隐式锁
小黄工程师学习进阶版
Javajava
隐式锁Java中的隐式锁(也称为内置锁或自动锁)是通过使用关键字实现的一种线程同步机制。当一个线程进入被synchronized修饰的方法或代码块时,它会自动获得对象级别的锁,退出该方法或代码块时则会自动释放这把锁。在Java中,隐式锁的实现机制主要包括以下两种类型:互斥锁(Mutex)虽然Java标准库并未直接暴露操作系统的互斥锁提供使用,但在Java虚拟机对synchronized关键字处理的
- Pillow 安装使用教程
小奇JAVA面试
安装使用教程pillowmicrosoft深度学习
一、Pillow简介Pillow是Python图像处理库PIL(PythonImagingLibrary)的友好分支,是图像处理的事实标准。它支持打开、编辑、转换、保存多种图像格式,常用于图像批量处理、验证码识别、缩略图生成等应用场景。二、安装Pillow2.1使用pip安装(推荐)pipinstallPillow2.2验证安装importPILprint(PIL.__version__)若无报错
- java 定时器需要守护线程,守护线程什么时候有用?
weixin_39956353
java定时器需要守护线程
当所有正在运行的线程都是守护进程线程时,JVM将退出。所以想象你正在写一个简单的游戏,你的主要方法循环,直到你决定退出。想象一下,在游戏开始的时候,你会开始一个无休止地轮询一些网站来触发警报的线程。当您决定结束游戏时,您希望JVM退出。你不希望无尽的投票,以防止游戏结束。所以你让这个轮询线程成为一个守护线程。当所有“正常”线程终止时,Deamon线程将被JVM自动终止。普通线程永远不会自动终止。您
- python炫酷烟花表白源代码-python炫酷烟花表白源代码
weixin_37988176
天天敲代码的朋友,有没有想过代码也可以变得很酷炫又浪漫?今天就教大家用Python模拟出绽放的烟花,工作之余也可以随时让程序为自己放一场烟花秀。python炫酷烟花表白源代码这个有趣的小项目并不复杂,只需一点可视化技巧,100余行Python代码和程序库Tkinter,最后我们就能达到下面这个效果:学完本教程后,你也能做出这样的烟花秀。整体概念梳理我们的整个理念比较简单。如上图示,我们这里通过让画
- PyTorch:Dropout 操作 torch.nn.Dropout()
sweettea~
Pythonpytorch深度学习
torch.nn.Dropout()是PyTorch中对Dropout层的其中一个实现,该函数底层调用torch.nn.functional.dropout();1、torch.nn.Dropout(p=0.5,inplace=False)其作用是,在training模式下,基于伯努利分布抽样,以概率p对张量input的值随机置0;training模式中,对输出以1/(1-p)进行scaling,
- java观察者模式
3213213333332132
java设计模式游戏观察者模式
观察者模式——顾名思义,就是一个对象观察另一个对象,当被观察的对象发生变化时,观察者也会跟着变化。
在日常中,我们配java环境变量时,设置一个JAVAHOME变量,这就是被观察者,使用了JAVAHOME变量的对象都是观察者,一旦JAVAHOME的路径改动,其他的也会跟着改动。
这样的例子很多,我想用小时候玩的老鹰捉小鸡游戏来简单的描绘观察者模式。
老鹰会变成观察者,母鸡和小鸡是
- TFS RESTful API 模拟上传测试
ronin47
TFS RESTful API 模拟上传测试。
细节参看这里:https://github.com/alibaba/nginx-tfs/blob/master/TFS_RESTful_API.markdown
模拟POST上传一个图片:
curl --data-binary @/opt/tfs.png http
- PHP常用设计模式单例, 工厂, 观察者, 责任链, 装饰, 策略,适配,桥接模式
dcj3sjt126com
设计模式PHP
// 多态, 在JAVA中是这样用的, 其实在PHP当中可以自然消除, 因为参数是动态的, 你传什么过来都可以, 不限制类型, 直接调用类的方法
abstract class Tiger {
public abstract function climb();
}
class XTiger extends Tiger {
public function climb()
- hibernate
171815164
Hibernate
main,save
Configuration conf =new Configuration().configure();
SessionFactory sf=conf.buildSessionFactory();
Session sess=sf.openSession();
Transaction tx=sess.beginTransaction();
News a=new
- Ant实例分析
g21121
ant
下面是一个Ant构建文件的实例,通过这个实例我们可以很清楚的理顺构建一个项目的顺序及依赖关系,从而编写出更加合理的构建文件。
下面是build.xml的代码:
<?xml version="1
- [简单]工作记录_接口返回405原因
53873039oycg
工作
最近调接口时候一直报错,错误信息是:
responseCode:405
responseMsg:Method Not Allowed
接口请求方式Post.
- 关于java.lang.ClassNotFoundException 和 java.lang.NoClassDefFoundError 的区别
程序员是怎么炼成的
真正完成类的加载工作是通过调用 defineClass来实现的;
而启动类的加载过程是通过调用 loadClass来实现的;
就是类加载器分为加载和定义
protected Class<?> findClass(String name) throws ClassNotFoundExcept
- JDBC学习笔记-JDBC详细的操作流程
aijuans
jdbc
所有的JDBC应用程序都具有下面的基本流程: 1、加载数据库驱动并建立到数据库的连接。 2、执行SQL语句。 3、处理结果。 4、从数据库断开连接释放资源。
下面我们就来仔细看一看每一个步骤:
其实按照上面所说每个阶段都可得单独拿出来写成一个独立的类方法文件。共别的应用来调用。
1、加载数据库驱动并建立到数据库的连接:
Html代码
St
- rome创建rss
antonyup_2006
tomcatcmsxmlstrutsOpera
引用
1.RSS标准
RSS标准比较混乱,主要有以下3个系列
RSS 0.9x / 2.0 : RSS技术诞生于1999年的网景公司(Netscape),其发布了一个0.9版本的规范。2001年,RSS技术标准的发展工作被Userland Software公司的戴夫 温那(Dave Winer)所接手。陆续发布了0.9x的系列版本。当W3C小组发布RSS 1.0后,Dave W
- html表格和表单基础
百合不是茶
html表格表单meta锚点
第一次用html来写东西,感觉压力山大,每次看见别人发的都是比较牛逼的 再看看自己什么都还不会,
html是一种标记语言,其实很简单都是固定的格式
_----------------------------------------表格和表单
表格是html的重要组成部分,表格用在body里面的
主要用法如下;
<table>
&
- ibatis如何传入完整的sql语句
bijian1013
javasqlibatis
ibatis如何传入完整的sql语句?进一步说,String str ="select * from test_table",我想把str传入ibatis中执行,是传递整条sql语句。
解决办法:
<
- 精通Oracle10编程SQL(14)开发动态SQL
bijian1013
oracle数据库plsql
/*
*开发动态SQL
*/
--使用EXECUTE IMMEDIATE处理DDL操作
CREATE OR REPLACE PROCEDURE drop_table(table_name varchar2)
is
sql_statement varchar2(100);
begin
sql_statement:='DROP TABLE '||table_name;
- 【Linux命令】Linux工作中常用命令
bit1129
linux命令
不断的总结工作中常用的Linux命令
1.查看端口被哪个进程占用
通过这个命令可以得到占用8085端口的进程号,然后通过ps -ef|grep 进程号得到进程的详细信息
netstat -anp | grep 8085
察看进程ID对应的进程占用的端口号
netstat -anp | grep 进程ID
&
- 优秀网站和文档收集
白糖_
网站
集成 Flex, Spring, Hibernate 构建应用程序
性能测试工具-JMeter
Hmtl5-IOCN网站
Oracle精简版教程网站
鸟哥的linux私房菜
Jetty中文文档
50个jquery必备代码片段
swfobject.js检测flash版本号工具
- angular.extend
boyitech
AngularJSangular.extendAngularJS API
angular.extend 复制src对象中的属性去dst对象中. 支持多个src对象. 如果你不想改变一个对象,你可以把dst设为空对象{}: var object = angular.extend({}, object1, object2). 注意: angular.extend不支持递归复制. 使用方法: angular.extend(dst, src); 参数:
- java-谷歌面试题-设计方便提取中数的数据结构
bylijinnan
java
网上找了一下这道题的解答,但都是提供思路,没有提供具体实现。其中使用大小堆这个思路看似简单,但实现起来要考虑很多。
以下分别用排序数组和大小堆来实现。
使用大小堆:
import java.util.Arrays;
public class MedianInHeap {
/**
* 题目:设计方便提取中数的数据结构
* 设计一个数据结构,其中包含两个函数,1.插
- ajaxFileUpload 针对 ie jquery 1.7+不能使用问题修复版本
Chen.H
ajaxFileUploadie6ie7ie8ie9
jQuery.extend({
handleError: function( s, xhr, status, e ) {
// If a local callback was specified, fire it
if ( s.error ) {
s.error.call( s.context || s, xhr, status, e );
}
- [机器人制造原则]机器人的电池和存储器必须可以替换
comsci
制造
机器人的身体随时随地可能被外来力量所破坏,但是如果机器人的存储器和电池可以更换,那么这个机器人的思维和记忆力就可以保存下来,即使身体受到伤害,在把存储器取下来安装到一个新的身体上之后,原有的性格和能力都可以继续维持.....
另外,如果一
- Oracle Multitable INSERT 的用法
daizj
oracle
转载Oracle笔记-Multitable INSERT 的用法
http://blog.chinaunix.net/uid-8504518-id-3310531.html
一、Insert基础用法
语法:
Insert Into 表名 (字段1,字段2,字段3...)
Values (值1,
- 专访黑客历史学家George Dyson
datamachine
on
20世纪最具威力的两项发明——核弹和计算机出自同一时代、同一群年青人。可是,与大名鼎鼎的曼哈顿计划(第二次世界大战中美国原子弹研究计划)相 比,计算机的起源显得默默无闻。出身计算机世家的历史学家George Dyson在其新书《图灵大教堂》(Turing’s Cathedral)中讲述了阿兰·图灵、约翰·冯·诺依曼等一帮子天才小子创造计算机及预见计算机未来
- 小学6年级英语单词背诵第一课
dcj3sjt126com
englishword
always 总是
rice 水稻,米饭
before 在...之前
live 生活,居住
usual 通常的
early 早的
begin 开始
month 月份
year 年
last 最后的
east 东方的
high 高的
far 远的
window 窗户
world 世界
than 比...更
- 在线IT教育和在线IT高端教育
dcj3sjt126com
教育
codecademy
http://www.codecademy.com codeschool
https://www.codeschool.com teamtreehouse
http://teamtreehouse.com lynda
http://www.lynda.com/ Coursera
https://www.coursera.
- Struts2 xml校验框架所定义的校验文件
蕃薯耀
Struts2 xml校验Struts2 xml校验框架Struts2校验
>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>>
蕃薯耀 2015年7月11日 15:54:59 星期六
http://fa
- mac下安装rar和unrar命令
hanqunfeng
mac
1.下载:http://www.rarlab.com/download.htm 选择
RAR 5.21 for Mac OS X 2.解压下载后的文件 tar -zxvf rarosx-5.2.1.tar 3.cd rar sudo install -c -o $USER unrar /bin #输入当前用户登录密码 sudo install -c -o $USER rar
- 三种将list转换为map的方法
jackyrong
list
在本文中,介绍三种将list转换为map的方法:
1) 传统方法
假设有某个类如下
class Movie {
private Integer rank;
private String description;
public Movie(Integer rank, String des
- 年轻程序员需要学习的5大经验
lampcy
工作PHP程序员
在过去的7年半时间里,我带过的软件实习生超过一打,也看到过数以百计的学生和毕业生的档案。我发现很多事情他们都需要学习。或许你会说,我说的不就是某种特定的技术、算法、数学,或者其他特定形式的知识吗?没错,这的确是需要学习的,但却并不是最重要的事情。他们需要学习的最重要的东西是“自我规范”。这些规范就是:尽可能地写出最简洁的代码;如果代码后期会因为改动而变得凌乱不堪就得重构;尽量删除没用的代码,并添加
- 评“女孩遭野蛮引产致终身不育 60万赔偿款1分未得”医腐深入骨髓
nannan408
先来看南方网的一则报道:
再正常不过的结婚、生子,对于29岁的郑畅来说,却是一个永远也无法实现的梦想。从2010年到2015年,从24岁到29岁,一张张新旧不一的诊断书记录了她病情的同时,也清晰地记下了她人生的悲哀。
粗暴手术让人发寒
2010年7月,在酒店做服务员的郑畅发现自己怀孕了,可男朋友却联系不上。在没有和家人商量的情况下,她决定堕胎。
12月5日,
- 使用jQuery为input输入框绑定回车键事件 VS 为a标签绑定click事件
Everyday都不同
jspinput回车键绑定clickenter
假设如题所示的事件为同一个,必须先把该js函数抽离出来,该函数定义了监听的处理:
function search() {
//监听函数略......
}
为input框绑定回车事件,当用户在文本框中输入搜索关键字时,按回车键,即可触发search():
//回车绑定
$(".search").keydown(fun
- EXT学习记录
tntxia
ext
1. 准备
(1) 官网:http://www.sencha.com/
里面有源代码和API文档下载。
EXT的域名已经从www.extjs.com改成了www.sencha.com ,但extjs这个域名会自动转到sencha上。
(2)帮助文档:
想要查看EXT的官方文档的话,可以去这里h
- mybatis3的mapper文件报Referenced file contains errors
xingguangsixian
mybatis
最近使用mybatis.3.1.0时无意中碰到一个问题:
The errors below were detected when validating the file "mybatis-3-mapper.dtd" via the file "account-mapper.xml". In most cases these errors can be d