
文献要点:
通过质谱流式检测单细胞间的不同染色质标记
细胞类型特异的染色质标记剖图预示免疫细胞的身份
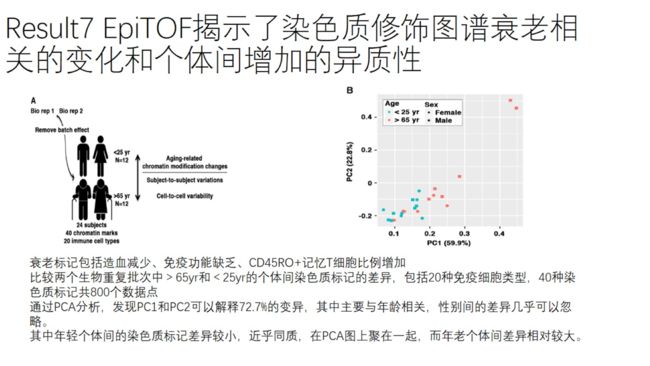
个体间和单细胞间的染色质变异随年龄增加而增加
染色质年龄相关的变化很大程度上是非遗传因素驱动





具体panel见文献的supplement
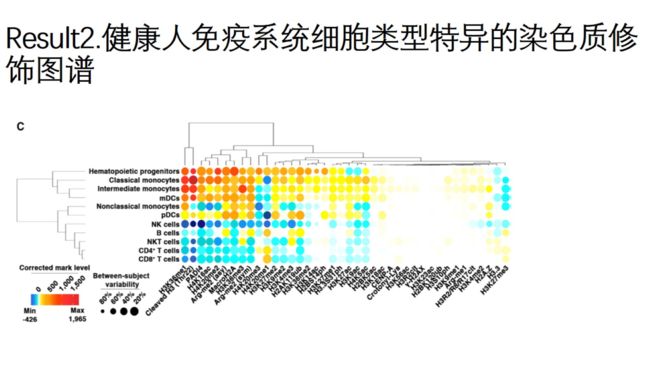
圆的直径代表多样性指数-逆辛普森指数(直径越大,可变性越小)
圆的颜色代表marker的平均表达水平
取样:PBMC
生物重复1:12个CMV-血清阴性的健康人,25岁以下6人,25岁以上6人,男女比例1:1
生物重复2:同上
线性回归模型,应用总组蛋白作为预测变量,对每种染色质修饰进行校正
校正后结果:
CD14+monocytes中cleaved H3比B、NK、T细胞含量高,瓜氨酸化酶PADI4也较高
反之,CD14+monocytes中H3.3、H3K27me3含量较低
CD4+和CD8+T细胞之间具有共同表达模式
NK细胞大部分的染色质marks的水平都比其他细胞类型低
r-H2AX和瓜氨酸化mark在未干扰的细胞中都呈低表达的状态
通过逆辛普森指数分析个体间的变异度,结果显示个体间的染色质修饰marks呈中等程度的变异度
结果显示,不同免疫细胞类型具有特定的染色质修饰图谱。
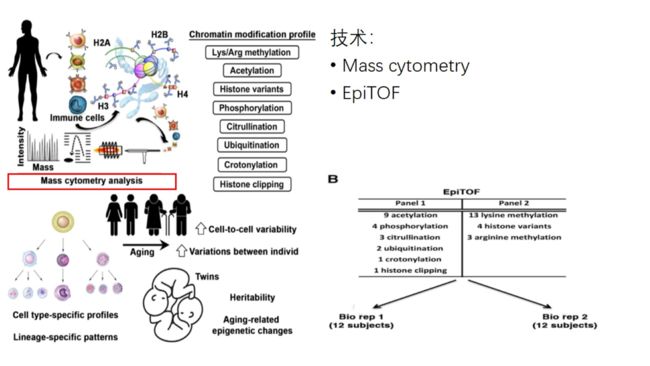

western blot对应蛋白的生物学意义:
Cleaved H3(Thr22):组织蛋白酶L对H3组蛋白进行酶解,去除染色质修饰标记
H3.3组蛋白变体
H3K27me3甲基化
PADI4 瓜氨酸化相关的酶-肽基精氨酸脱亚酶4
H3R2cit瓜氨酸化
r-H2AX磷酸化
H4对照
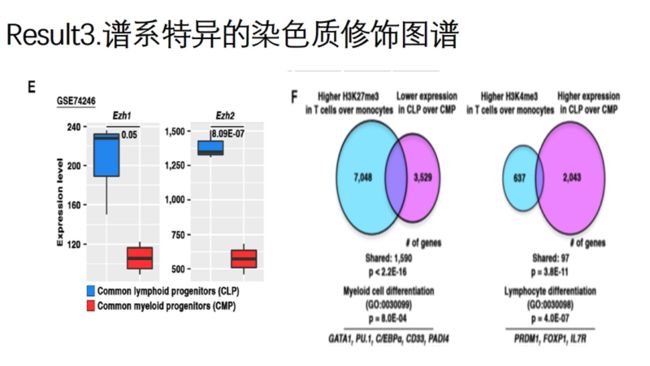
EZH1/EZH2:生成H3K27me3的赖氨酸甲基转移酶
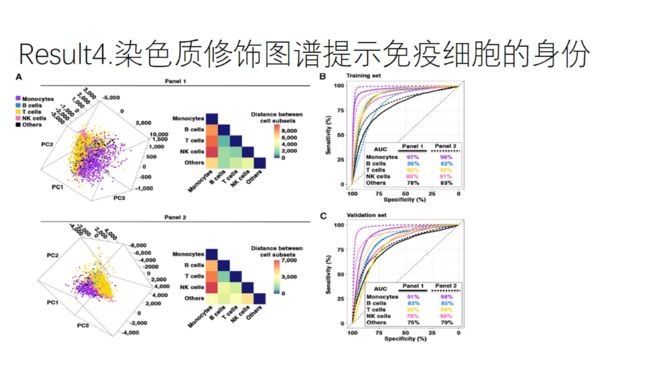
1.PCA分析
根据单细胞EpiTOF的数据进行PCA分析,细胞聚类结果如图A:说明同一类型的细胞具有相同的染色质修饰模式
计算每种细胞类型之间的欧氏距离,发现monocytes和其他几种淋巴细胞的距离最大,反映了淋巴系和髓系细胞的不同分化过程。
淋巴细胞之间的距离远低于淋巴细胞与monocytes之间距离的1/4.
2.回归模型:L-1 regularized logistic regressionmodels
AUC曲线评价
图B:将bio rep1作为训练集,利用单细胞的染色质修饰的数据对免疫细胞进行分类;无论用哪个panel,都能准确的将NK、B、T细胞、monocytes从PBMC中分辨出来。
图C:将bio rep2作为验证集,利用bio rep1的系数可以分辨出相对应的细胞类型。
说明了利用染色质修饰可以对免疫细胞的身份进行鉴定。
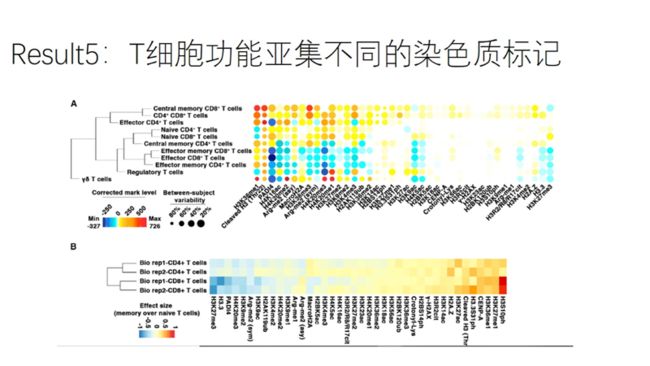
构建质朴流式针对T细胞的EpiToF的panel,可以分析11种T细胞亚集的40种染色质标记
图A:在上述队列针对T细胞的染色质标记进行分析
表达αβ-TCR的T细胞的染色质修饰标记与表达γδ-TCR的T细胞不同,使其在免疫功能方面也有所不同
CD4+和CD8+nativeT细胞的染色质标记图谱基本一致
CD45RO-效应细胞和CD45RO+记忆细胞的染色质标记开始出现分化
说明T细胞染色质标记的可塑性,随着在适应性免疫过程中细胞分化为确保特异性免疫功能而被重新配置。
图B:两个bio rep的关联分析,提示较高的生物可重复性
nativeT细胞分化为memoryT细胞时与其染色质修饰相关。在CD4+和CD8+T细胞中发现同样的趋势。
记忆T细胞中,H3S10的磷酸化水平升高,这一修饰通常在高转录基因的转录起始部位发现;同时发现常染色质相关的H3K27me1和H3K36me1增高,异染色质相关的H3K27me3, H4K20me3和H3K9me2低表达,说明记忆T细胞的染色质比native T细胞的染色质处于更开放的状态


PC1的评分的双峰分布与MixTool基于CD56的分离模式结果类似。
CD56dimNK细胞占NK细胞的90%,具有更强的杀伤细胞的功能;CD56brightNK细胞则是分泌细胞因子的主要来源。
在CD56bringhtNK细胞中发现更多乙酰化标记,PADI4和cleaved histone H3 Thr22两个髓系特异标志物也高表达,重新分析EpiTOF的数据发现, CD56bringhtNK细胞不与CD56dimNK细胞和其他淋巴细胞聚类在一起。
另有新的研究表明,两种NK细胞可能分化来源于不同的造血谱系。
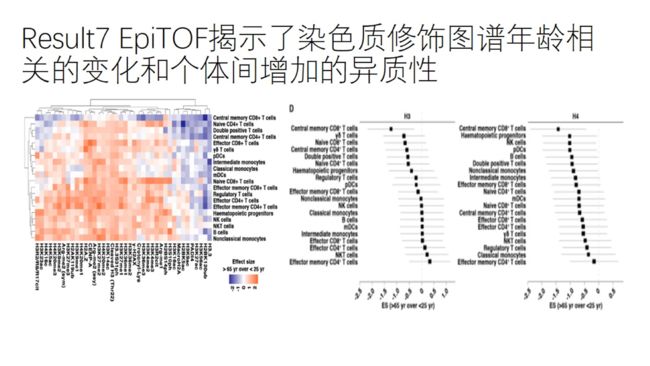
衰老相关的染色质标记的增加并非组蛋白总量的增加,因为细胞水平上的H3和H4含量未有明显差异;
但我们共同观察到,随着衰老,不同免疫细胞类型的染色质标记都呈现出增加的趋势。
但中心记忆性CD8+T细胞中,34个染色质标记呈现出下降的趋势,同时伴随着H3和H4组蛋白明显下降。
之前研究表明,在几个物种发现了衰老相关的组蛋白的丢失;以及衰老相关的核小体占有率下降与染色质的动力学和转录活性密切相关。
证实了中心记忆性CD8+T细胞中染色质的独特的衰老模式。

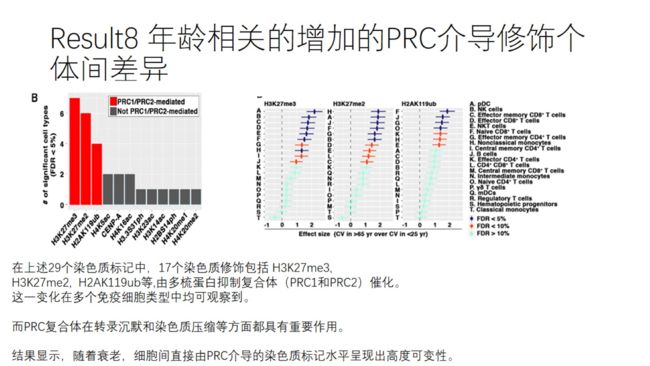


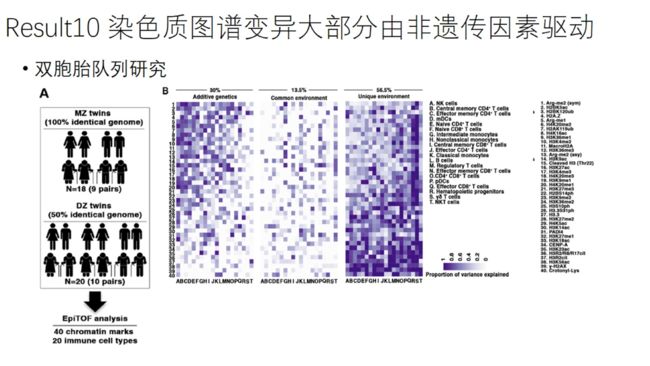
双胞胎队列研究:纳入9对同卵双胞胎 10对异卵双胞胎
经典ACE模型:A:附加遗传 C:共同环境 E:独特环境
环境因素解释了70%的染色质图谱变异,遗传因素的作用也并非不重要的(30%)
CD45RO-CCR7+naive,CD45RO+CCD7+ central memory 、CD45RO+CCD7 effector memory CD4+ T cells占CD4+T细胞的比例主要受遗传因素的影响
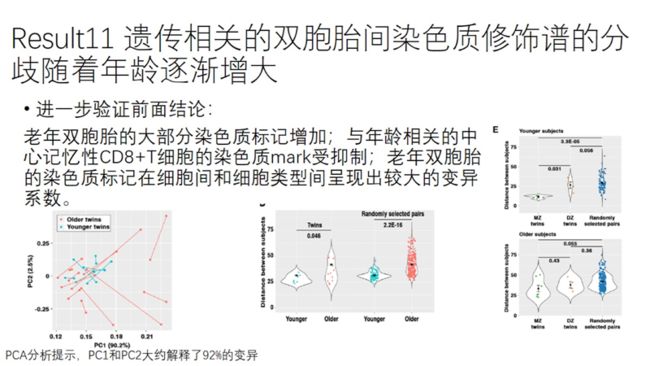
比较老年双胞胎间和随机筛选的遗传上不相关的个体间的欧氏距离,老年个体明显高于年轻个体,说明染色质修饰谱随年龄增加的变异主要由环境因素所驱动。
年轻个体间,MZ双胞胎的染色质图谱相较DZ双胞胎和随机配对组的更为一致,说明了附加遗传和表观遗传决定早年染色质图谱的重要性。
但是年老个体间,MZ、DZ和随机配对组的在PCA上的欧氏距离并不是差异很大,也说明了环境因素和后天累积的体细胞突变对染色质修饰谱的重要影响。




小结:(个人的观点)
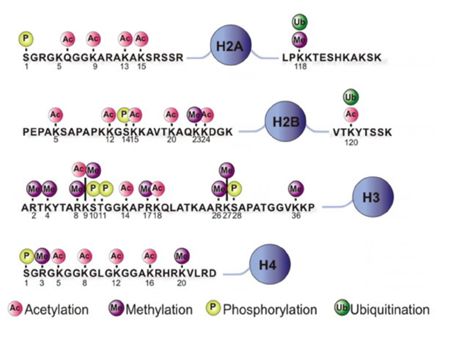
首先,这篇文献虽然题目说了是单细胞体系的研究,但用的不是现在如火朝天的严格意义的单细胞测序技术如10xgenomics,BD等,采用的是质谱流式技术,通过流式的原理将细胞分为单细胞体系。质谱流式技术是利用已有经典的染色质标记蛋白对应的抗体和金属离子标记作为label先入为主将细胞分类,不同于10x等通过转录组或全基因组测序等然后无监督或者半监督聚类分析,在根据各类细胞的marker表达情况或者基因表达模式定义各个细胞群的类型。
文献研究表观组学的主要技术是EpiTOF,表观飞行时间,可以认为是蛋白组学发展而来的技术,只是这里的蛋白主要是研究染色质上的组蛋白及其修饰后的蛋白。
质谱流式结合EpiTOF这一技术的一个缺点是和细胞流式分析一样,需要所研究的蛋白相应的抗体,如果没有相应抗体就没办法分析。
我们知道的表观遗传学其实包括很多方面:
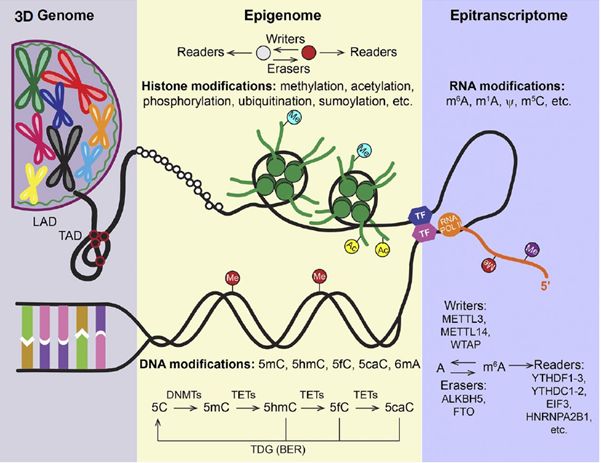
像18年底10x公司出的ATAC-seq技术,则是研究全染色质的开放状态,像这样的技术还有DNase-seq、FAIRE-seq、MNase-seq等;
17年北大汤富酬教授开发的single-cell COOL-seq实现了单细胞表观多组学测序,在单细胞水平上同时进行5个层面上的分析包括染色质状态、DNA甲基化、基因组拷贝数变异、以及染色体倍性的全基因组测序技术等。
这篇文献则是从染色质中的组蛋白修饰状态这一角度进行分析,提供了不一样的表观组学的研究思路。