Effective leadership and decision-making in animal groups on the move.《动物团体在运动中的有效领导和决策》
Couzin, Iain D. , et al. “Effective leadership and decision-making in animal groups on the move.” Nature (London) 433.7025(2005):513-516.
文章目录
- 方法 Methods
- Group size
- Group direction
- Elongation
- Uncertainty of information
- Starting conditions
对于成群觅食或旅行的动物,做出运动决定通常取决于群体成员之间的社交互动。但是,在许多情况下,很少有人拥有相关信息,例如有关食物来源或迁移路线。使用一个简单的模型,我们展示了信息如何在组内传递,既不需要发送信号,也不需要成员知道哪些个体(如果有的话)拥有信息的情况下进行传递信息。我们发现,小组越大,指导小组所需的有识之士的比例就越小,并且只需要很小一部分的有识之士即可达到很高的准确性。我们还演示了群体如何做出一致的决定,即使知情的个人不知道他们是多数派还是少数派,与其他人相比他们的信息质量如何,甚至不知道是否有其他知情的个人。我们的模型为生物系统的有效领导和决策机制提供了新的见解。
关于群体中信息传递机制的主要问题包括:未被告知的个体如何识别那些被告知的个体,这种识别是否真的有必要,以及当被告知的个体偏好(即期望的运动方向)不同时,群体如何做出集体决策。据了解,一些动物物种已经进化出了特定的补充信号来帮助引导同种动物。在这方面最著名的是蜜蜂的摇摆舞,它招募蜂群成员去寻找食物来源。此外,有价值的经验可能与年龄或优势有关,这可能是由某些物种的同种特性来估计的。然而,当考虑到鱼群、有蹄类动物、昆虫和鸟类的迁徙时,这种解释是否站住脚仍然值得怀疑,因为它们的拥挤限制了个体间相互探测的范围。例如,在远洋鱼群中,个体之间的距离通常不到一个体长。尽管一些物种很可能有一个大方向由基因决定迁移倾向,或响应非生物因素,如热梯度可能援助的迁移,很可能对许多物种经历了小组成员发挥重要作用在指导那些缺乏经验或经验不足。在鱼群中,已知能够影响群体觅食行为和鱼群导航能力的知情个体相对较少。同样地,蜂群中很少有个体(大约5%)能引导蜂群到一个新的蜂巢。
此外,对于一些动物群体,如大型昆虫群或鱼群,假设群体成员具有个体识别能力可能是不合理的。在这里,我们讨论两个基本问题,都发生在缺乏复杂的信号机制的情况下,以及当组成员无法确定谁有信息,谁没有信息时。首先,如何在组内传输关于资源位置或迁移路线的信息;第二,当知情人士的偏好不同时,个体如何达成共识。
我们考虑了将个体分组的能力,即根据当地邻居的行为(社会互动)来调整他们的行为。群体由 N N N个个体组成。每个个体具有位置向量 c i ( t ) c_i(t) ci(t)、方向向量 v i ( t ) v_i(t) vi(t)和速度 s i s_i si,都试图始终保持自身与其他 j j j之间的最小距离 α \alpha α通过远离该范围内的邻居
d i ( t + Δ t ) = − ∑ j ≠ i c j ( t ) − c i ( t ) ∣ c j ( t ) − c i ( t ) ∣ (1) d_i(t+\Delta{t}) = -\sum\limits_{j\ne i}{\frac{{{c}_{j}}(t)-{{c}_{i}}(t)}{\left| {{c}_{j}}(t)-{{c}_{i}}(t) \right|}} \tag{1} di(t+Δt)=−j=i∑∣cj(t)−ci(t)∣cj(t)−ci(t)(1)
其中KaTeX parse error: Expected group after '_' at position 3: di_̲表示所需的行进方向。这模拟了个人为了保持个人空间和避免碰撞而采取的行动。逃避是最重要的。如果在这个区域内没有检测到邻居,那么个体将倾向于被局部相互作用范围 ρ \rho ρ内与 j j j个邻居所吸引和与其保持一致:
d i ( t + Δ t ) = ∑ j ≠ i c j ( t ) − c i ( t ) ∣ c j ( t ) − c i ( t ) ∣ + ∑ j = 1 v j ( t ) ∣ v j ( t ) ∣ (2) d_i(t+\Delta{t}) = \sum\limits_{j\ne i}{\frac{{{c}_{j}}(t)-{{c}_{i}}(t)}{\left| {{c}_{j}}(t)-{{c}_{i}}(t) \right|}} + \sum\limits_{j = 1}{\frac{{{v}_{j}}(t)}{\left| {{v}_{j}}(t) \right|}} \tag{2} di(t+Δt)=j=i∑∣cj(t)−ci(t)∣cj(t)−ci(t)+j=1∑∣vj(t)∣vj(t)(2)
这里将 d i ( t + Δ t ) d_i(t+\Delta{t}) di(t+Δt)转换为单位向量 d ^ i ( t + Δ t ) = d i ( t + Δ t ) / ∣ d i ( t + Δ t ) ∣ {{\hat{d}}_{i}}(t+\Delta t)={{d}_{i}}(t+\Delta t)/|{{d}_{i}}(t+\Delta t)| d^i(t+Δt)=di(t+Δt)/∣di(t+Δt)∣。
为了探究具有先验知识的成员对群体的影响, p p p的一部分被给予了关于首选方向的信息(模拟为单位向量g),例如,表示到已知资源的方向,或者迁移路线的一部分。所有其他的个体都是无知的,没有期望的、特定的移动方向,也不知道组内哪些个体有信息,哪些没有。知情个体通过加权项 ω \omega ω,并使用 d ′ i ( t + Δ t ) {{{d}'}_{i}}(t+\Delta t) d′i(t+Δt)代替 d ^ i ( t + Δ t ) {{\hat{d}}_{i}}(t+\Delta t) d^i(t+Δt)来衡量他们偏好的方向对集群的影响;其中
d ′ i ( t + Δ t ) = d ^ i ( t + Δ t ) + ω g i ∣ d ^ i ( t + Δ t ) + ω g i ∣ (3) {{{d}'}_{i}}(t+\Delta t)=\frac{{{{\hat{d}}}_{i}}(t+\Delta t)+\omega {{g}_{i}}}{\left| {{{\hat{d}}}_{i}}(t+\Delta t)+\omega {{g}_{i}} \right|} \tag{3} d′i(t+Δt)=∣∣∣d^i(t+Δt)+ωgi∣∣∣d^i(t+Δt)+ωgi(3)
如果 ω = 0 \omega=0 ω=0,则向量 g i g_i gi没有影响,个体没有意愿向任何特定方向移动。当 ω \omega ω接近1时,个体倾向于平衡他们向 g i g_i gi方向移动的偏好和他们与群体成员保持社会互动的愿望。当 ω \omega ω大于1时,个体受其偏好的方向 g i g_i gi的影响比受其邻居的影响更大(有关模型的详细信息,请参见方法部分)。
组的精度,即信息传递的质量,可以量化为组方向(参见方法)围绕首选方向 g g g的归一化角偏差,最小值为0,最大值为1。
对于给定的群体规模,我们发现随着知情个体比例的增加,群体运动(在偏好方向上)的准确性逐渐增加。此外,随着群体规模的扩大,这种关系变得越来越非线性(图1a),这意味着群体越大,需要以给定的准确性指导群体的知情人士比例就越小。因此,对于足够大的群体,只需要非常小比例的知情个人即可接近最大的准确性。对于动物群体,如迁徙的蜜蜂群体,其成本随着的侦察兵(知道期望的方向的个体)的增加而增加,因为其通过通过摇尾舞吸引其他成员,以及为了让侦察兵学习基本的导航技能,均需要一定的时间。因此,我们可能期望蜂群已经进化即它们已经有效地达到精确的渐近线(图1a),并且我们预测它们将在拥有更多的侦察蜂种群中获得很少的好处。
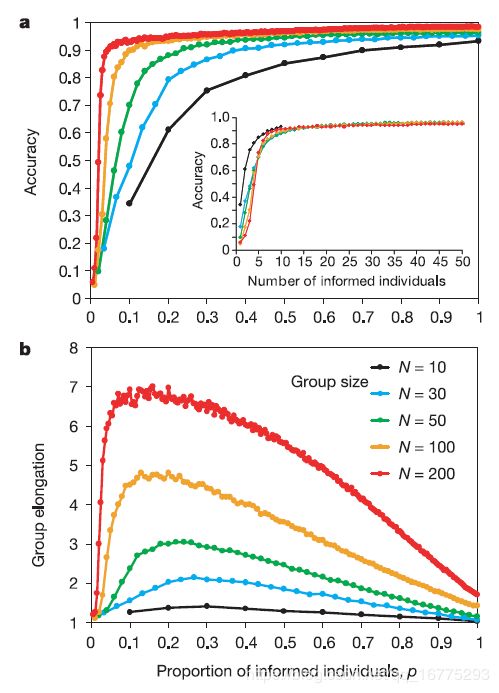
图 1 对不同的群体大小 N N N的群体而言,群体精度和形状可以看做是知情个体的比例 p p p的函数。随着 p p p的增加,图(a)中最初准确性的增加与群体随着边长有关。知情的个体倾向于在群体中占据一个前沿位置。随着 p p p的进一步增加,群体的延伸率随之下降,并且越来越多的群体知道方向向量 g g g. 400次重复实验, ω = 0.5 \omega=0.5 ω=0.5; α = 1 \alpha=1 α=1, ρ = 6 \rho=6 ρ=6, γ = 0 \gamma=0 γ=0, Δ t = 0.2 s \Delta{t}=0.2s Δt=0.2s, θ = 2 \theta=2 θ=2, s i = α S − 1 s_i=\alpha S^{-1} si=αS−1。
无论知情个体比例是大是小,首选方向 ω \omega ω的权重影响最不重要(图2a, c, d)。然而,在中间值时, ω \omega ω与群体准确性呈强正相关(图2b)。这对应于图1中精度增长率达到其渐近线的区域。然而,对于 p p p的这些中间值,增加 ω \omega ω是有代价的,因为在群体运动的准确性和群体碎片化的概率之间存在权衡(图2b)。缺乏群体凝聚力显然会对蜜蜂或鱼群造成不利影响,而且有实验证据表明,在其他动物群体中也会出现这种权衡,在这些动物群体中,群体对路线的学习取决于知情个体与无知个体保持社会互动的能力。

图2 首选方向加权的影响。群体运动的准确性(黑是圆圈)和群体破碎的可能性(红色三角形)随权重 ω \omega ω和知情个体占据的比例 p p p的变化而变化。图 a,p = 0.02(1个人); 图b,p = 0.1(5个人);图 c,p = 0.2(10个人);图 d,p = 0.5(25个人)。参数如图1所示,N = 50。
然而,由于经验或动机的不同,组内消息灵通的个体可能在他们喜欢的方向上有所不同。成群的动物经常不得不集体决定,比如一起移动到一个特定的资源,比如一个巢穴或食物来源。做出这样的决定的方法知之甚少,,特别是在大的群体的情况下,以及当个人不知道自己占多数还是少数,或者与其他人相比他们的信息质量(准确度)如何,或者他们的信息的质量与别人相比,甚至是否有任何其他个人拥有信息。为了研究这个问题,我们创建了两个知情个体的子集,每个子集都有自己的方向偏好。
如果表现出每个偏好的个体数量相等,则群体运动的方向取决于偏好偏好的差异程度:随着这种差异的增加,群体将以同样的概率的从有知情个体的偏好方向(一共两个偏好方向)中随机选择一个(图3A,a)。但是,如果表现出一种偏爱的个体数量增加(图3A,b和c),则该群体(在偏爱方面存在适当的差异,图3A,a)将共同选择多数人所偏爱的方向,即使该多数很小(图3A,b)。
一个群体在差异相对较小的情况下,群体对偏好进行平均,而在差异较大的情况下对多数选择达成共识的能力,可能在许多动物群体中都很重要,尤其是在可以调整这种转变发生的时间点的情况下。实现这一优化我们介绍了一个简单的反馈加权 ω \omega ω的首选方向。如果在一个给定的时间步长中,有先验知识的个体发现自己朝着自己的首选方向相似的方向(这里是在20度范围内)移动, ω \omega ω则会变大(由由 ω i n c \omega_{inc} ωinc,最大到, ω m a x \omega_{max} ωmax),否则降低(由 ω d e c \omega_{dec} ωdec,最低为0),这种反馈回路允许针对首选方向上的较小差异(图3B,C)达成共识性决策(与平均相反)。图3B,C显示,在每个子集中的个体数量发生微小变化时,就会发生向共识决策的转变,并且随着子集大小差异的增加,少数子集的影响会迅速减小。

图3 当知情的个人偏好不同时,群体方向的选择。每个组内包含知情个体的两个子集(分别具有方向优先级 s 1 s1 s1和 s 2 s2 s2,大小分别为 n 1 n1 n1和 n 2 n2 n2)的组的组方向的归一化概率分布(最大比例),每个组都有自己的方向性: s 1 = 0 s1=0 s1=0度(白色虚线), s 2 = 0 − 180 s2=0-180 s2=0−180度(2000次重复试验,每次以10度为间隔)。群体总大小为 N = 100 N=100 N=100(图1中的参数; ω \omega ω, ω m a x = 0.4 \omega_{max}=0.4 ωmax=0.4)。我们展示了稍微改变n1和n2对组方向的巨大影响。在图a中,第一列n1=n2(A,B=5; C=10,表明在更大的 p p p值下具有一致性)。在b中,第二列n1增加1,而在c中,第3列,n1增加1,n2减少1(以便与第一列直接比较)。 B和C包括反馈。 图A,a,n1=5,n2=5; A,b,n1=6,n2=5; A,c,n1=6,n2=4; B,ac,n1和n2分别与A,ac相同。 C,a,n1=10,n2=10; C,b,n1=11,n2=10. C,c,n1=11,n2=9.对于B和C, ω i n c = 0.012 \omega_{inc}=0.012 ωinc=0.012, ω d e c = 0.0008 \omega_{dec}=0.0008 ωdec=0.0008。白色实线仅供参考,代表所有知情个体的平均向量 g g g的方向(其中 ω \omega ω为常数)。
这种决策机制还允许在信息质量方面进行区分。例如,如果知情个体的两个子集的大小没有差异,但是他们正确确定其首选方向的能力存在差异,则该小组可以共同选择与最小错误相关的方向(图4,有关详细信息,请参见方法)。
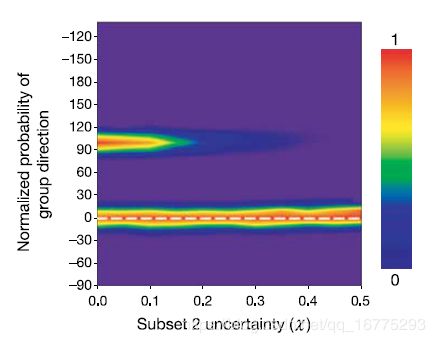
图4 根据信息质量(x,请参见方法)区分方向(s1 = 0; s2 = 100度)。 参数如图3所示。n1= 10,n2 =10。当x> 0时,子集2的信息不确定。
我们的模型表明,在没有明确信号或复杂信息传递机制的情况下,动物群内可以发生信息的有效传递和决策。这意味着知情和无知的人不必彼此识别,领导力可以根据成员之间的信息差异而出现,因此可以转移。尽管这些属性也可能影响群体运动,但无需调用个体之间的固有差异(例如,由于体型较大导致的优势)。此外,我们在此提出的协调机制只需要有限的认知能力,并表明个人可以对具有信息的人自发做出反应。这对于我们对群体觅食,社交学习,迁移和导航的理解很重要,并且可能为在分组机器人之间进行信息传递提供新的设计协议。
方法 Methods
动物的运动受到随机影响(例如,感觉/运动错误)。 这是通过修改 d ′ i ( t + Δ t ) {{{d}'}_{i}}(t+\Delta t) d′i(t+Δt)或 d ^ i ( t + Δ t ) {{{\hat{d}}}_{i}}(t+\Delta t) d^i(t+Δt)来模拟的; 对于无知和知情的个体,分别将其旋转一个以圆形包裹的高斯分布为中心的随机角(以0为中心),标准差为 σ = 0.01 \sigma=0.01 σ=0.01弧度(请参见下面的“信息不确定性Uncertainty of information”),从而得出向量 d ′ ′ i ( t + Δ t ) {{{d}''}_{i}}(t+\Delta t) d′′i(t+Δt) :每个人在时间 Δ t \Delta t Δt内最多可以其渴望的方向转动 θ Δ t \theta{\Delta t} θΔt弧度; 如果转动的角度,即 v i ( t ) v_i(t) vi(t)与 d ′ ′ i ( t + Δ t ) {{{d}''}_{i}}(t+\Delta t) d′′i(t+Δt)之间的夹角小于 θ Δ t \theta{\Delta t} θΔt,则它们与所需矢量对齐,即 v i ( t + Δ t ) = d ′ ′ i ( t + Δ t ) {{v}_{i}}(t+\Delta t)={{{d}''}_{i}}(t+\Delta t) vi(t+Δt)=d′′i(t+Δt); 否则,他们会向该方向转动 θ Δ t \theta{\Delta t} θΔt角度。 个体 i i i的新位置矢量可以通过 c i ( t + Δ t ) = c i ( t ) + v i ( t + Δ t ) Δ t s i {{c}_{i}}(t+\Delta t)={{c}_{i}}(t)+{{v}_{i}}(t+\Delta t)\Delta t{{s}_{i}} ci(t+Δt)=ci(t)+vi(t+Δt)Δtsi计算得到,其中 s i s_i si是个体 i i i的速度。
Group size
这里的群体大小可与鱼群、鸡群、畜群或其他群体的大小相比,但比诸如蜜蜂殖民地的大型聚群体要小,这是因为随着群体大小N的增加,计算机处理时间也会非线性增加。 但是,我们的结果可能会在保持团体成员凝聚力的约束下,独立于绝对团体规模(即与群体大小无关)。 为了测试群体是否保持凝聚力,我们使用了参考文献18、19中所述的等效类技术。
Group direction
为了量化群体的方向 h h h,我们创建了一个向量,该向量表示群体质心(the group’s centroid)在时间 t f Δ t − 50 Δ t t_f\Delta{t}-50\Delta{t} tfΔt−50Δt到时间 t f Δ t t_f\Delta_{t} tfΔt之间的延伸,其中 t f t_f tf表示最后的时间步长,为2500。 在图1和2中,我们计算了400个重复实验的角度平均偏差(the mean angular deviation) s s s,相当于计算了线性标准偏差(the linear standard deviation),我们对其进行了归一化处理,所以该偏差最小值为0,对应于无信息传递(即群体沿随机方向移动);其最大值为1,对应模拟群体的运动方向始终与 g g g完全相同。
Elongation
通过在与行进方向对齐的群体周围创建边界框并计算与群体方向对齐的轴的长度与垂直于群体方向的轴的长度之比,来测量伸长率。 当两个轴相同时,此值为1;当该组在行进方向上变长时,该值为>1;当该组在垂直于行进方向时变长时,该值为<1。
Uncertainty of information
个体可能还没有完全了解其渴望的方向 g g g,这可以在仿真开始时通过对每个个体 i i i旋转相同类型的具有标准偏差 γ \gamma γ的圆环高斯分布进行模拟,从而得出相应的向量 g i g_i gi。 改变 γ \gamma γ可以在群体能够保持凝聚力所施加的上限内定量地而非定性地改变我们的结果(参见补充图1)。
为了f仿真同一群体内具有先验知识的不同个体,正确确定其期望的方向的能力差异,将 p / 2 p/2 p/2个个体(s2)的 g g g以标准差为 χ \chi χ的高斯分布角,标准差x弧度(创建 g i g_i gi),在仿真开始时(参见图4)。
Starting conditions
仿真中每个个体都从随机的位置和随机的方向开始的。