问题:
linkage disequilibrium (LD)和 pairwise correlation的区别?似乎它们都能达到相同的目的。
先从直觉上理解一下GWAS的原理:
核心就是SNP与表型的关联,对于每一个genome位点,如果某个SNP总是与某疾病同时出现,那我们就可以推测这个SNP极有可能会导致这个疾病。
规范点讲就是看某个SNP在case和control两个population间是否有allel frequency的显著差异。
而现实情况是,我们样本数有限,而且有时候control和case样本不平衡,样本还分男女、人群,而且我们需要对3亿个碱基位点都做统计检验。
我们应该设计哪些指标来评价一个snp与表型的关联呢?
思考:如果一个位点有多个SNP,而只有其中的一个SNP与疾病相关怎么办?
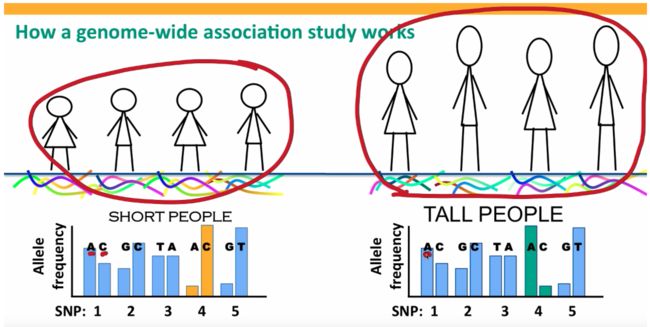
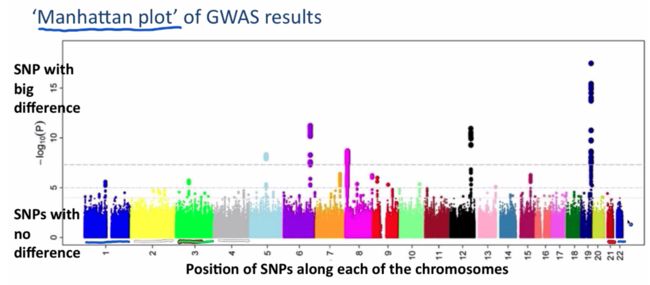
牢记:曼哈顿图中的点代表的不是样品,而是SNP。
思考:曼哈顿图中,显著的SNP并不是鹤立鸡群的冒出来,而是似乎被捧出来的,就像高楼大厦一样,从底下逐步冒出来的。这一座大厦其实就是连锁在一起的SNP,具有很高的LD score。
思考:虽然曼哈顿图里每个点是SNP,但是通常都会把最显著的SNP指向某个基因,因为大家最关注的还是SNP的致病根源,但这样找出来的只有编码区的SNP。
注意:最突出的SNP极有可能不是causal SNP,它只是near the causal SNP。问题就来了,怎么找causal SNP呢?fine mapping
基本背景
什么是SNP?进化过程中随机产生的单点突变,并能稳定的在群体中遗传。
什么是allele frequency in population?每一个genome位点都有两个或多个allele,不同allel之间有明显的频率上的差异,简单点理解就是A和a两个性质的频率,但这里是碱基位点,而不是性状基因。
GWAS分析的前提
sample size足够,学过统计的都知道sample size会影响power,没有足够的power是得不出正确结论的,GWAS通常需要大量的样本,几千是标配,几百就太少,现在有的都达到了几万几十万级别;
一个大误区就是GWAS会测全基因组WGS,其实不是的,那太贵了,大部分是做DNA chip DNA芯片,只包含了常见的10^6个SNP。稍微有钱的就会上WES,就会得到所有编码区的SNP;最有钱的就是WGS了,全部检测,编码非编码,常见罕见,1000genome就是靠这个才NB的。
大致原理已经讲了,其实还有统计原理,暂时略过,先看实操。
怎么用PLINK来做GWAS?油管视频:GWAS in Plink 里面有paper、示例数据、代码下载,可以跑跑熟悉一下。
发表了paper的,GWAS pipeline:A tutorial on conducting genome‐wide association studies: Quality control and statistical analysis。
github地址
一下着重讲解一下这个流程的操作细节:
主要是四方面的分析:
- All essential GWAS QC steps along with scripts for data visualization.
- Dealing with population stratification, using 1000 genomes as a reference.
- Association analyses of GWAS data.
- Polygenic risk score (PRS) analyses.
先看下PLINK的文本文件格式:
ped:行是个体,列是表型和SNP的基因型数据;
map:snp的特征数据;
二进制有三个格式:
主要就是把ped拆成了fam和bed,map变成了bim。
通常要做covariate分析,所以还有个covariate文件。

QC:
| Step | Command | Function |
|---|---|---|
| 1: Missingness of SNPs and individuals | ‐‐geno | Excludes SNPs that are missing in a large proportion of the subjects. In this step, SNPs with low genotype calls are removed. |
| ‐‐mind | Excludes individuals who have high rates of genotype missingness. In this step, individual with low genotype calls are removed. | |
| 2: Sex discrepancy | ‐‐check‐sex | Checks for discrepancies between sex of the individuals recorded in the dataset and their sex based on X chromosome heterozygosity/homozygosity rates. |
| 3: Minor allele frequency (MAF) | ‐‐maf | Includes only SNPs above the set MAF threshold. |
| 4: Hardy–Weinberg equilibrium (HWE) | ‐‐hwe | Excludes markers which deviate from Hardy–Weinberg equilibrium. |
| 5: Heterozygosity | For an example script see https://github.com/MareesAT/GWA_tutorial/ | Excludes individuals with high or low heterozygosity rates |
| 6: Relatedness | ‐‐genome | Calculates identity by descent (IBD) of all sample pairs. |
| ‐‐min | Sets threshold and creates a list of individuals with relatedness above the chosen threshold. Meaning that subjects who are related at, for example, pi‐hat >0.2 (i.e., second degree relatives) can be detected. | |
| 7: Population stratification | ‐‐genome | Calculates identity by descent (IBD) of all sample pairs. |
| ‐‐cluster ‐‐mds‐plot k | Produces a k‐dimensional representation of any substructure in the data, based on IBS. |
fine mapping
一个常识就是GWAS是2007年才出现得,所以2017年才出了篇有名的综述ten years of GWAS,fine mapping是GWAS后才出现得。
实验室很早就开始研究fine mapping了:2009 - Fine mapping of the 9q31 Hirschsprung’s disease locus
看一下introduction,什么是fine mapping?
目的很简单:GWAS找到的大多不是causal variants,fine mapping就是就fill这个gap。
GWAS得到大体的SNP后,必须做两方面的深入分析:
第一步就是对SNP给一个概率上的causality,这就是fine-mapping;第二步就是根据功能注释来确定该SNP确实能导致某个基因。
The first is to assign well-calibrated probabilities of causality to candidate variants, known as fine-mapping. The second step is to try to connect these variants to likely genes whose perturbation leads to altered disease risk by functional annotation.
基本原理:
Strategies for fine-mapping complex traits -
Integrative modeling of eQTLs and cis-regulatory elements suggests mechanisms underlying cell type specificity of eQTLs
Although eQTLs are increasingly used to provide mechanistic interpretations for human disease associations, the cell type specificity of eQTLs presents a problem. Because the cell type from which a given physiological phenotype arises may not be known, and because eQTL data exist for a limited number of cell types, it is critical to quantify and understand the mechanisms generating cell type specific eQTLs. For example, if a GWAS identifies a set of SNPs associated with risk of type II diabetes, the researcher must choose a target cell type to develop a mechanistic model of the molecular phenotype that causes the gross physiological change. One can imagine that the relevant cell type might be adipose tissue, liver, pancreas, or another hormone-regulating tissue. Furthermore, if the GWAS SNP produces a molecular phenotype (i.e., is an eQTL) in lymphoblastoid cell lines (LCLs), it is not necessarily the case that the SNP will generate a similar molecular phenotype in the cell type of interest. Furthermore, there are many examples of cell types with particular relevance to common diseases, for example dopaminergic neurons and Parkinson's disease, that lack comprehensive eQTL data or catalogs of CREs. The utility of eQTLs for complex trait interpretation will therefore be improved by a more thorough annotation of their cell type specificity.
eQTL最大的问题还是celltype的特异性不够,关键还是要celltype的定义足够精准!
现在GWAS已经属于比较古老的技术了,主要是碰到严重的瓶颈了,单纯的snp与表现的关联已经不够,需要具体的生物学解释,这些snp是如何具体导致疾病的发生的。
而且,大多数病找到的都不是个别显著的snp,大多数都找到了很多的snp,而且snp都落在非编码区了,这就导致对这些snp的解读非常的困难。
GWAS的核心结果就两个,曼哈顿图和QQ-plot,看懂就够了。
单纯会跑GWAS pipeline已经没什么价值了,现在重在下游的分析,有几个热点:
- Polygenic risk score (PRS) analyses
- meta-analysis
The International HapMap Project (http://hapmap. ncbi.nlm.nih.gov/; Gibbs et al., 2003) described the patterns of com- mon SNPs within the human DNA sequence whereas the 1000 Genomes (1KG) project (http://www.1000genomes.org/; Altshuler et al., 2012) provided a map of both common and rare SNPs.
common和rare就是根据allele frequency来界定的,但是似乎没有明确界限。
HapMap用的是array,所有测得都是一些人为挑的点,所以就是common snps;而1000 genomes是WGS,所以包含了所有的点,所以有common和rare一起。
GWAS和核心就是LD,目前大部分的GWAS都是测得array,因为便宜。
GWAS会漏掉很多点,所以才会有fine-mapping,根据haplotype来做一些imputation。
Linkage disequilibrium (LD)连锁不平衡:不同基因座位的各等位基因在人群中以一定的频率出现。在某一群体中,不同座位某两个等位基因出现在同一条染色体上的频率高于预期的随机频率的现象。(就是孟德尔的分离不是随机的,在染色体上越靠近的allele越倾向于绑在一起,属于物质性的限制。)
例如两个相邻的基因A B, 他们各自的等位基因为a b. 假设A B相互独立遗传,则后代群体中观察得到的单倍体基因型 AB 中出现的P(AB)的概率为 P(A) * P(B). 实际观察得到群体中单倍体基因型 AB 同时出现的概率为P(AB)。 计算这种不平衡的方法为: D = P(AB)- P(A) * P(B).
事实上,可以检测遍布基因组中的大量遗传标记位点snp,或者候选基因附近的遗传标记来寻找到因为与致病位点距离足够近而表现出与疾病相关的位点,这就是等位基因关联分析或连锁不平衡定位基因的基本思想。
待看的paper:Strategies for fine-mapping complex traits
assign well-calibrated probabilities of causality to candidate variants, known as fine-mapping.
还有一些非常重要的概念:
effect size:效应量
power:功效,power analyses
Underestimated Effect Sizes in GWAS: Fundamental Limitations of Single SNP Analysis for Dichotomous Phenotypes
在语境里理解:One explanation of the missing heritability is that complex diseases are caused by a large number of causal variants with small effect sizes.
PRS combines the effect sizes of multiple SNPs into a single aggregated score that can be used to predict disease risk
haplotype phasing单倍体分型
Positions with 00 and 11 are called homozygous positions. Positions with 10 or 01 are called heterozygous positions. We note that the reference genome is neither the paternal nor the maternal genome but the genome of an un-related human (or more precisely the mixture of genomes of a few individuals). An individual’s haplotype is the set of variations in that individual’s chromosomes. We note that as any two human haplotypes are 99.9% similar, the mapping problem can be solved quite easily.
Haplotype phasing is the problem of inferring information about an individual’s haplotype. To solve this problem, there are many methods.
Lecture 10: Haplotype Phasing - Community Recovery
参考:PLINK | File format reference
vcftools
plink的主要功能:数据处理,质量控制的基本统计,群体分层分析,单位点的基本关联分析,家系数据的传递不平衡检验,多点连锁分析,单倍体关联分析,拷贝数变异分析,Meta分析等等。
首先必须了解plink的三种格式:bed、fam和bim。(注意:这里的bed和我们genome里的区域文件bed完全不同)
plink需要的格式一般可以从vcf文件转化而来 (顺便了解一下ped和map两种格式):
PED: Original standard text format for sample pedigree information and genotype calls. Normally must be accompanied by a .map file. 谱系信息和基因型信息。每一行是一个人。
MAP: Variant information file accompanying a .ped text pedigree + genotype table. 变异信息。每一行是一个变异 | snp。
# PED
1 1 0 0 1 0 G G 2 2 C C
1 2 0 0 1 0 A A 0 0 A C
1 3 1 2 1 2 0 0 1 2 A C
2 1 0 0 1 0 A A 2 2 0 0
2 2 0 0 1 2 A A 2 2 0 0
2 3 1 2 1 2 A A 2 2 A A
# MAP
1 snp1 0 1
1 snp2 0 2
1 snp3 0 3# vcf转ped和map
plink --vcf file.vcf --recode --out file
# ped和map转bed、bim和fam
plink --file test --make-bed --out test
三种格式的官方介绍
bed文件(真实的bed文件是二进制的,比较难读)
bed:Primary representation of genotype calls at biallelic variants. Must be accompanied by .bim and .fam files. Loaded with --bfile; generated in many situations, most notably when the --make-bed command is used. Do not confuse this with the UCSC Genome Browser's BED format, which is totally different. 基因型信息。所以转换后就是一个matrix,每一行是一个个体,每一列就是一个变异。其中0、1、2分别对应了aa、Aa或aA和AA。不考虑碱基型,因为我们不关注ATGC的变化。
fam:Sample information file accompanying a .bed binary genotype table. 样本信息。每一行就是一个样本。
bim:Extended variant information file accompanying a .bed binary genotype table. 每一行是一个变异,及其注释信息。
rs4970383 rs3748592 rs9442373 rs1571150 rs6687029
2431:NA19916 2 0 0 0 1
2424:NA19835 1 0 1 2 0
2469:NA20282 1 0 1 0 1
2368:NA19703 0 0 0 2 0
2425:NA19901 1 0 1 2 2OR
# xxd -b test.bed
00000000: 01101100 00011011 00000001 11011100 00001111 11100111 l.....
00000006: 00001111 01101011 00000001 .k.
- First two bytes 01101100 00011011 for PLINK v1.00 BED file
- Third byte is 00000001 (SNP-major) or 00000000 (individual-major)
- Genotype data, either in SNP-major or individual-major order
- New "row" always starts a new byte
- Each byte encodes up to 4 genotypes
- 10 indicates missing genotype, otherwise 0 and 1 point to allele 1 or allele 2 in the BIM file, respectively
- Bits in each byte read in reverse order
fam文件
1 2431 NA19916 0 0 1
2 2424 NA19835 0 0 2
3 2469 NA20282 0 0 2
4 2368 NA19703 0 0 1
5 2425 NA19901 0 0 2OR
1 1 0 0 1 0
1 2 0 0 1 0
1 3 1 2 1 2
2 1 0 0 1 0
2 2 0 0 1 2
2 3 1 2 1 2
bim文件
1 1 rs4970383 0 828418 A
2 1 rs3748592 0 870101 A
3 1 rs9442373 0 1052501 C
4 1 rs1571150 0 1464167 A
5 1 rs6687029 0 1508931 COR
1 snp1 0 1 G A
1 snp2 0 2 1 2
1 snp3 0 3 A C
基本概念
关联分析:就是AS的中文,全称是GWAS。应用基因组中数以百万计的单核苷酸多态;SNP为分子遗传标记,进行全基因组水平上的对照分析或相关性分析,通过比较发现影响复杂性状的基因变异的一种新策略。在全基因组范围内选择遗传变异进行基因分析,比较异常和对照组之间每个遗传变异及其频率的差异,统计分析每个变异与目标性状之间的关联性大小,选出最相关的遗传变异进行验证,并根据验证结果最终确认其与目标性状之间的相关性。
连锁不平衡:LD,P(AB)= P(A)*P(B)。不连锁就独立,如果不存在连锁不平衡——相互独立,随机组合,实际观察到的群体中单倍体基因型 A和B 同时出现的概率。P (AB) = D + P (A) * P (B) 。D是表示两位点间LD程度值。
曼哈顿图:在生物和统计学上,做频率统计、突变分布、GWAS关联分析的时候,我们经常会看到一些非常漂亮的manhattan plot,能够对候选位点的分布和数值一目了然。位点坐标和pvalue。map文件至少包含三列——染色体号,SNP名字,SNP物理位置。assoc文件包含SNP名字和pvalue。haploview即可画出。
CMplot:一个R包,画曼哈顿图的。
q-q plot:分位数-分位数图,assess if a set of data plausibly came from some theoretical distribution such as a Normal or exponential.
GCAT(Genome-wide Complex Trait Analysis):在分析的时候计算LD,PCA以及关联分析。
BLUP:即最佳线性无偏预测(Best Linear Unbiased Prediction),该方法广泛用于GWAS中对多年多点表型数据分析当中,R语言中的lme4包可以对此进行分析。
常识:世界范围的人类群体,在表型上可谓千差万别,但是基因组上的差异却非常小,而且这种差异大多数表现为SNP (Single nucleotide polymorphism , 单核苷酸多态性)。
IBS:在两个或两个以上的个体当中,如果一个DNA片段具有相同的核苷酸序列,就说这个DNA片段是IBS。
IBD: 如果IBS片段是遗传自同一个祖先且中间过程没有发生过重组事件,就说这个片段是IBD。
数据表示模型:由1 和2 组成的2n个序列,每一个SNP 基因型对应两个序列。对于任意一个个体的SNP 基因型数据进行处理(忽略ACGT 的差别)如22,21,12,11 分别对应于SNP 基因型,aa aA Aa AA。然后把这些序列转换为 由0、1、2 组成的数量为n的SNP 序列,表示为:

第i个个体的SNP基因型为:

这两个个体间的第K个snp的IBS状态为:


个体i和个体j的SNP的IBS 状态值非0的区域满足一定阈值就作为候选IBD片段,可以表示为:

把N个体的数据分成case和control两组进行分析,其中case包含个l个体,control包含m个个体,然后对这两组数据分别进行评价分析,对每个SNP 得到各自的S值。差异值最大的snp位点就可能为我们的候选位点。

一些问题
这些文件中的0,1,2是什么意思?
跑跑PLINK工具
plink --bfile --pheno --pheno-name t16 --linear hide-covar --covar --covar-name
AGE,SEX,PC1,PC2,PC3,PC4 --ci 0.95 --out
--bfile 将snp文件变成二进制格式
--pheno 这里导入我们刚刚处理的性状文件
--pheno-name t16 要处理的性状名字是t16
--linear hide-covar 使用线性模型,hide-covar指的是不要对我没加入的协变量进行分析
--covar --covar-name AGE,SEX,PC1,PC2,PC3,PC4 把我们选取的协变量加入线性回归模型中,我们选的协变量有:AGE,SEX,PC1,PC2,PC3,PC4
--ci 0.95 设置置信区间
SNP过滤问题
使用vcftools过滤:
1. MAF<0.05
vcftools --vcf test.vcf --maf 0.05 --out XX
2.完整度大于90%
vcftools --vcf test.vcf --max-missing 0.9 --OUT XX
3.平均深度大于5
vcftools --vcf test.vc --min-meanDP 5 --out xx
注:
使用--gvcf更为快捷
使用plink过滤
1.vcf转化plink格式
vcftools --vcf test.vcf --plink --out xxx
2.plink --noweb --file plink --geno 0.05 --maf 0.05 --hwe 0.0001 --make-bed
跟一个官网的教学,无需写代码,教学材料:Resources available for download 非常通俗,容易入门。
ped文件:谱系信息和基因型;
Contains no header line, and one line per sample with 2V+6 fields where V is the number of variants. The first six fields are the same as those in a .fam file.
The seventh and eighth fields are allele calls for the first variant in the .map file ('0' = no call); the 9th and 10th are allele calls for the second variant; and so on.
前6行就和fam文件一样,家庭id,家庭内id,性别,表型。
后面两个一组,比如第7和第8就是map中第一个snp的等位基因(人有两条染色体,每条DNA都是双链的,不考虑双链,因为有互补配对)。
fam文件:样本信息;
- Family ID ('FID')
- Within-family ID ('IID'; cannot be '0')
- Within-family ID of father ('0' if father isn't in dataset)
- Within-family ID of mother ('0' if mother isn't in dataset)
- Sex code ('1' = male, '2' = female, '0' = unknown)
- Phenotype value ('1' = control, '2' = case, '-9'/'0'/non-numeric = missing data if case/control)
map文件:突变信息;
- Chromosome code. PLINK 1.9 also permits contig names here, but most older programs do not.
- Variant identifier
- Position in morgans or centimorgans (optional; also safe to use dummy value of '0')
- Base-pair coordinate
bim文件:额外的突变信息;
- Chromosome code (either an integer, or 'X'/'Y'/'XY'/'MT'; '0' indicates unknown) or name
- Variant identifier
- Position in morgans or centimorgans (safe to use dummy value of '0')
- Base-pair coordinate (normally 1-based, but 0 ok; limited to 231-2)
- Allele 1 (corresponding to clear bits in .bed; usually minor)
- Allele 2 (corresponding to set bits in .bed; usually major)
MAF, Minor allele frequency: SNPs with a minor allele frequency of 0.05 or greater were targeted by the HapMap project. 最小等位基因频率
QC
The SNPs are currently coded according to NCBI build 36 coordinates on the forward strand.
Data quality control in genetic case-control association studies
plink可以对snp进行QC过滤,根据一些指标,比如MAF。。。
plink的结果必须要有了解,
1. 将文本的ped和map文件转化为二进制的bed、bim和fam文件;
2. 关联分析的结果,其实就是给每个人赋值一个表型,然后就做关联分析,得到每一个snp与表型的相关性,用p-value来表示,最终可以画曼哈顿图;
CHR SNP BP A1 F_A F_U A2 CHISQ P OR 1 rs3094315 792429 G 0.1489 0.08537 A 1.684 0.1944 1.875 1 rs4040617 819185 G 0.1354 0.08537 A 1.111 0.2919 1.678 1 rs4075116 1043552 C 0.04167 0.07317 T 0.8278 0.3629 0.5507 1 rs9442385 1137258 T 0.3723 0.4268 G 0.5428 0.4613 0.7966
参考:
利用PLINK进行GWAS分析(一)
GWAS的基本原理 讲得比较通俗
QQ plot图——评价你的统计模型是否合理 讲得比较清楚
基于全基因组snp数据如何进行主成分分析(PCA)- GCTA
2019年02月16日更新