The Plant Journal | DAP-seq助力揭示乌龙茶树杂种优势形成的分子机制

期刊:The Plant Journal
影响因子:7.1
发表日期:2022年10月11日
单位:中国农业科学院深圳农业基因组研究所
2022年10月11日中国农业科学院深圳农业基因组研究所合成生物学研究中心张兴坦研究员课题组在The Plant Journal上发表了题为“Allele-specific expression and chromatin accessibility contribute to heterosis in tea plants (Camellia sinensis)”的研究论文。该研究基于亲本基因组对茶树杂交种进行了等位基因鉴定和偏倚分析。作者利用ATAC-seq和DAP-seq联合分析,揭示了茶树杂种的等位基因和染色质的调控机制,为茶树杂种优势的形成和调控提供新的思路。爱基百客为该研究提供DAP-seq的技术支持。
研究背景
茶树是一种多年生木本植物,富含复杂而有特色的次生代谢产物。茶树是重要的经济作物,通过特定的加工或制造方法,可以制成各种口味的饮料,受到许多消费者的喜爱。茶树具有高度杂合性,不同亲本包含两套不同的染色体,导致F1杂种性状高度分离。从茶叶育种的角度来看,只有具有杂种优势的茶树才能被优先选择。杂种优势指的是一种常见的生物现象,即杂交品种优于亲本,并依赖亲本系间的互补遗传变异。目前,优良茶树品种的育种已有多年研究,许多杂交品种得到广泛栽培,带来了巨大的经济价值。然而,关于茶叶杂种优势形成的分子机制仍是空白。因此,本研究主要利用DAP-seq与ATAC-seq联合分析来揭示乌龙茶树杂种优势形成的分子机制。
材料和方法
研究材料:乌龙茶树(亲本:铁观音,黄丹;杂交种:金观音,黄观音)
测序策略:ATAC-seq、DAP-seq和RNA-seq
研究思路
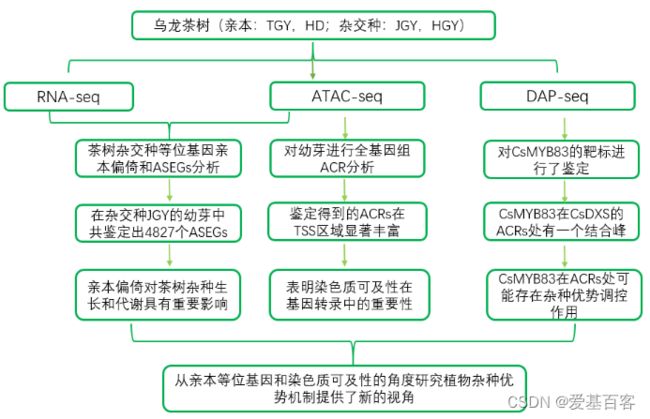
研究结果
1. 茶树杂交种等位基因亲本偏倚和ASEGs分析
为探讨茶树杂种优势中等位基因亲本偏倚和染色质可及性的调控,作者对几种茶树进行了全基因组RNA-seq和ATAC-seq分析,结果表明亲本对茶树杂交种等位基因的表达有重要影响。为了进一步研究茶树杂交种等位基因的亲本偏倚,作者系统地鉴定了杂交种的等位基因特异性表达基因(ASEGs)。在杂交种JGY的幼芽中共鉴定出4827个ASEGs, 2250个偏母系TGY表达,2577个偏父系HD表达(图1c),HGY也表现出一致的亲本偏倚。这些结果表明,大多数ASEGs具有保守的亲本偏倚表达。此外,作者还对杂种中具有不同亲本偏差的ASEGs进行了KEGG富集分析。结果表明亲本偏倚对茶树杂种生长和代谢具有重要影响。

图1. 茶树杂交种亲本偏倚等位基因和ASEGs的全局分析
2. 在茶树杂种中具有亲本偏倚的ASEGs有助于挥发性萜类物质的积累
萜类是茶树的重要代谢类,它们对茶树的生物和非生物胁迫的抗性和茶叶的香气有重要影响。因此,作者研究了相应的亲本等位基因对它们的调控机制。茶树萜烯的合成有两条典型途径:胞质甲戊酸(MVA)和质体磷酸甲基赤糖醇(MEP)途径(图2a)。MVA和MEP通路中分别有9个和8个ASEGs,它们分别负责茶树中重要的倍半萜和单萜的合成(图2b)。结果显示,显性亲本等位基因的高表达有利于增加萜类的积累,MVA和MEP通路上游1-2拷贝的结构基因大多受不同亲本偏倚的调控,亲本等位基因的显性表达有利于萜类的部分或完全显性效应。
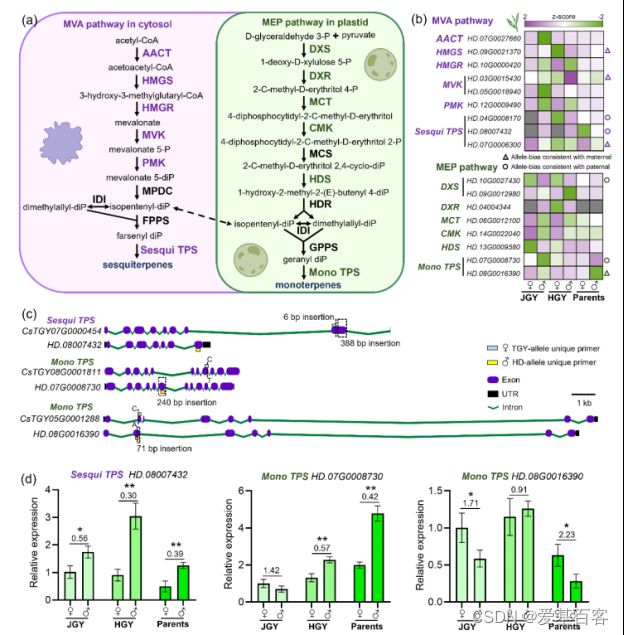
图2.MVA和MEP通路中的ASEGs
3. 具有亲本偏倚的ASEGs参与调节嘌呤化合物的杂种优势
接下来,作者还研究了嘌呤代谢,它不仅对植物生命至关重要,还涉及到茶树特有的优质代谢产物的合成。杂交种JGY和HGY在嘌呤代谢中表现出杂种优势,在作者之前的研究中,它与不同亲本的遗传密切相关。作者在嘌呤代谢途径中鉴定出15个ASEGs(图3c),它们可能参与嘌呤化合物不同亲本遗传和杂种优势的形成。作为嘌呤代谢途径起始的核心基因,核磷酸焦磷酸激酶HD等位基因(CsPRPS)的高表达可能是HGY积累更多嘌呤化合物的重要因素,而腺苷激酶TGY等位基因(CsAK)的高表达可能会导致JGY中ADP含量丰富。此外,5'-核苷酸酶合成的黄嘌呤(Cs5'-NT)是嘌呤代谢和嘌呤生物碱合成途径的重要分支,它们在JGY的两个亲本等位基因中均有较高的表达,这可能是导致嘌呤生物碱含量较高的原因。考虑到磷酸核糖焦磷酸激酶、5 '核苷酸酶和腺苷酸激酶的重要性,作者根据它们的等位基因变异设计了3个特异性引物集(图3d)。作者观察到与转录组结果一致的亲本偏差(图3e),这说明显性等位基因可能是后续嘌呤代谢物导向育种的重要目标。
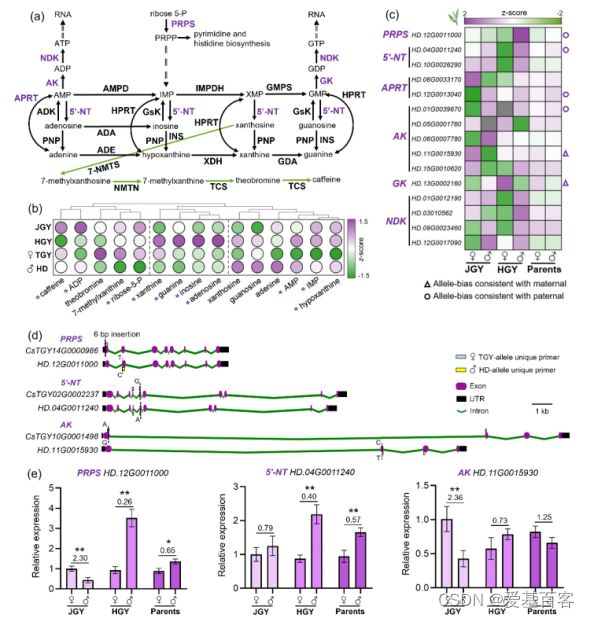
图3. 嘌呤和嘌呤生物碱代谢途径中的ASEGs分析
4. 利用ATAC-seq分析茶杂种及其亲本染色质可及性特征
为了研究茶树杂交种和亲本之间的调控景观,作者利用ATAC-seq对它们的幼芽进行全基因组可及染色质区域(ACR)分析。在重复性高的茶杂交种及其亲本中,共鉴定出17591-29513个ACRs,这些ACRs在转录起始位点(TSS)区域显著丰富,并显示出一个尖峰(图4a)。茶树杂交种和亲本的ACR总序列长度为7.66 (HD) ~ 11.65 (JGY) Mb,约占总基因组大小的0.26 ~ 0.40%。杂交种的ACR数量和总序列长度均显著大于亲本,表明杂交种基因组染色质可及性更强。然后,作者进一步研究了ACRs在茶树基因组中的分布。结果显示,茶树杂交种在所有基因组位置上都比亲本表现出更多的ACRs,尤其是dACRs(基因间的远端ACRs)和启动子pACRs (TSS上游2 kb内的近端ACRs和TTS下游2 kb内的远端ACRs) (图4c)。茶树杂交种和亲本共有10950 ~ 18229个dACR,占ACR总数的61.02-62.06%,远高于其他类型的ACR。此外,随着染色质可及性水平的提高,其相应基因的表达也会发生相应的变化,这表明染色质可及性在基因转录中的重要性(图4f)。
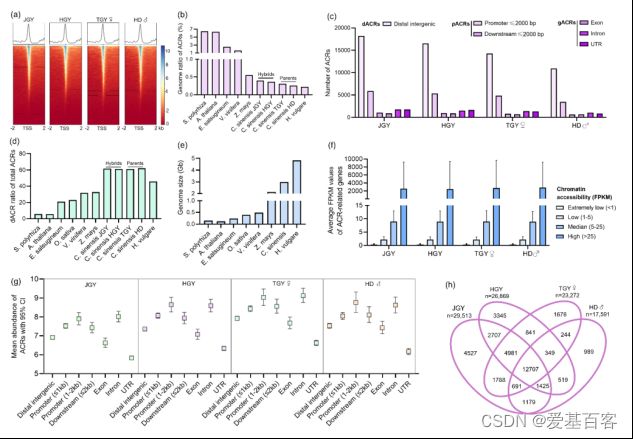
图4. 茶树杂交种及其亲本染色质可及性分析
5. 杂交特异性ACR的分析和重复出现的motif的鉴定
为了进一步分析杂交种特异性ACR的相关基因,作者对ATAC-seq结果进行进一步分析。KEGG途径富集分析结果证明了杂交种特异性ACR相关基因使杂交种在某些代谢通路上可以对杂种优势的调控有促进作用。接下来,作者用MEME-ChIP进一步研究杂交特异性ACR中重复出现的转录因子(TF)结合motif (p值≤0.05)(图5c)。结果显示,在杂交种JGY和HGY的特异性ACRs中分别发现了5个高频出现的TF结合motif,这些与植物的生长代谢和激素合成有关,它们对ACR位置分布的偏好也相似,这说明在优良的茶树杂种中生长和代谢杂种优势的形成具有共同的特定调控作用。
此外,作者还发现在蓝光下调节叶片发育和生物钟的TCP22,以及长下胚轴5 (HY5)结合的motif在JGY特异性ACRs中高频出现;而调控拟南芥耐盐性的NAC57和调控叶缘和花瓣发育的TCP5,在HGY中高频出现,这些反映了茶树杂交种各自不同的调控网络。

图5. 茶树中杂交特异性ACR和重复出现的motif
6. 茶树杂种优势转录调控网络的构建与差异可及ACRs 的Motif鉴定
为了进一步研究调控网络,通过评估杂交与亲本之间不同组间的差异ACRs,作者确定了两个杂交品种相对于不同亲本的差异ACRs相关基因。结果显示,可及性上调的ACR相关基因在杂种中数量总是远远高于亲本 (图6a),这反映了杂种中染色质的正向调控,它可能有助于杂种的生长和杂种优势的形成。
此外,为了评估染色质可及性水平的差异对相应基因丰度的影响,作者还分析了TGY与JGY和与HGY中ACR相关差异基因集的表达(图6b)。结果表明了杂种和亲本的基因表达水平受相应染色质可及性的调节。KEGG分析结果表明,茶树杂交种中可及性上调的ACR相关基因主要参与以甘氨酸、丝氨酸和苏氨酸代谢为核心的能量和次生代谢调控网络,这说明杂种中相对亲本可及性更高的ACR对某些代谢物杂种优势形成的调控作用。
最后,考虑到杂交种和亲本之间的差异ACRs代表着或多或少可及的染色质区域,可能包含重要的顺式调控转录因子,作者还通过MEME-ChIP识别出差异ACRs的过度表达的顺式调控基序,在杂交种可及性增加的ACRs中发现了6个显著富集的TF结合motif(图6d),这6个转录因子可能在调节茶杂种优势形成中起主导作用。此外,作者在ACRs中获得了3个过表达的TF结合基元,它们在杂交中可及性显著降低(图6d),其中包括一个调节碳和氮代谢的Dof1.4型因子,一个TCP成员TCP9调控靶基因与茉莉酸、水杨酸、气孔发育和非生物胁迫响应相关和MYB93,后者负责调控侧根发育,并影响苯丙素途径和次脂素沉积。在杂交种可及性降低的ACRs中,MYB93调控的靶点最多,并与Dof1.4和TCP9协同调控部分ACRs的可及性水平(图6f),从而控制茶杂交种的部分代谢途径。

图6. 杂交种和亲本之间ACR相关基因和等位基因及其转录调控网络的可及性差异
7. 染色质可及性在茶树杂种两种正杂种优势代谢途径中对ASEGs的作用
为了研究染色质可及性对具有正杂种优势的代谢途径中ASEGs的调节作用,作者分别在17和15个萜类和嘌呤类生物碱代谢途径的ASEGs中鉴定出8和16个ACR。嘌呤和嘌呤生物碱代谢途径中的ASEGs表现出明显高于萜类代谢途径的ACR频率和可及水平(图7a)。在嘌呤核苷酸和生物碱代谢途径中,杂交HGY中ACRs的平均丰度高于其他茶树品种。此外,作者还选取茶杂交种萜烯合成途径中可及水平较高的3个ACRs在IGV浏览器上进行可视化研究,结果显示,这三个ACRs包含多个潜在的MYB、ARF和NAC结合位点 (图7d)。据报道,这些靶点的多个同源TF成员参与了萜类合成途径中许多结构基因的激活,并影响萜类的含量。为了研究这些靶点,作者利用DAP-seq技术,对CsMYB83的靶标进行了鉴定。结果表明,CsMYB83在CsDXS的ACRs处有一个结合峰(图7d),这说明CsMYB83在ACRs处可能存在杂种优势调控作用。
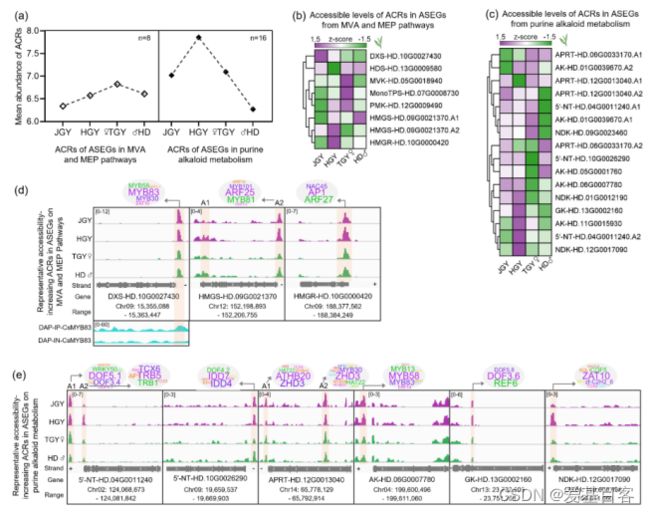
图7. ASEGs中来自MVA、MEP和嘌呤代谢途径的ACRs分析
◆ 小 结 ◆
在本研究中,作者对中国栽培面积最大的人工乌龙茶进行了等位基因和染色质可及性水平分析。作者首先找到了杂种优势形成的原因,然后利用ATAC-seq发现了杂交种的ACRs明显高于亲本,这可能使杂交种的基因转录活性更广泛、更强。杂交种中显著增加的ACR有助于调节ASEG的表达,从而影响杂种优势代谢产物的形成。此外,作者还通过DAP-seq验证了速率限制酶CsDXS启动子ACR中CsMYB85的结合motif。这些结果为从亲本等位基因和染色质可及性的角度研究植物杂种优势机制提供了新的视角。然而,该研究无法识别等位基因特异性ACR并分析其与ASEGs的关系。因此,后续研究可以通过构建多个杂交种的单倍型解析基因组,更精确地探索杂种优势形成的等位基因调控。
文章链接:https://doi.org/10.1111/tpj.16004
◆ DAP-Seq简介 ◆
DAP-Seq(DNA affinity purification sequencing)是一种高通量筛选TF结合位点的方法,实验中使用体外表达的TF蛋白对裸露的全基因组DNA片段进行孵育,以确定结合序列。DAP-seq将蛋白质体外表达技术与高通量测序技术相结合,不需要针对每个转录因子制备特异性抗体,所以DAP-seq具有快速、高通量、节约时间成本等显著优势。
爱基百客在DAP-seq上拥有丰富的项目经验,涉及物种包括:
植物:番茄、葡萄、龙眼、大豆、小麦、水稻、苦荞、棉花、茶树、杉木、三叶青、白菜等;
动物:鲤鱼、斜纹夜蛾。
有相关需求老师可以联系我们哦!