文献阅读(一)植物应对干旱的生理学反应 | The physiology of plant responses to drought
分享一篇Science上的综述文章,主要探讨了植物应对干旱的生理机制,强调通过调控激素信号提升植物耐旱性、保障粮食安全的重要性。
摘要
干旱每年致使农作物产量的损失,比所有病原体造成损失的总和还要多。为适应土壤中的湿度梯度变化,植物会改变自身生理机能,调整根系的生长和结构,并关闭地上部分的气孔。这些组织特异性反应改变了细胞信号的传导,致使植物提前开花或生长不良,且往往会造成产量下降。对模式植物拟南芥进行的生理学和分子分析显示,植物激素信号传导是调节植物对干旱或缺水反应的关键。在本文中,我们将探讨如何通过调控特定细胞和细胞区域的激素信号,来促进植物更好地应对干旱。我们还会探究目前已有的知识,以及在培育高产、抗旱作物这一研究方向上,未来需要解决的核心问题。。
重要概念
植物抗旱性:植物感知缺水信号并启动应对策略的这种能力。
植物抗旱性通过多种机制发挥作用:
- 逃避机制(escape):在可能阻碍其生存的干旱胁迫出现之前,植物加速进入生殖阶段。这是一种在干旱造成严重损害前,完成繁殖过程以保证物种延续的策略。比如一些沙漠植物,会在短暂的降雨期迅速开花结果,赶在干旱加剧前完成繁殖。
- 避免机制(avoidance):植物通过增加内部含水量并防止组织受损来忍受干旱。这可以通过多种方式实现,例如增强根系对水分的吸收能力,减少水分从叶片表面的散失(如关闭气孔),以及在细胞内积累一些能够调节渗透压的物质,使植物细胞保持水分,从而避免干旱对组织造成伤害。
- 耐受机制(tolerance):即使内部含水量较低,植物仍能在干旱期间维持生长。植物会启动一系列生理和生化过程来适应低水状态,例如激活抗氧化系统清除干旱胁迫产生的有害活性氧物质,维持细胞内的代谢平衡,调整自身的生长和发育模式,以在缺水条件下尽可能地维持生命活动。
在细胞层面,干旱信号会促进脯氨酸和海藻糖等应激保护代谢物的生成,触发抗氧化系统以维持氧化还原稳态,并调动过氧化物酶来防止细胞受到急性损伤,维持细胞膜的完整性。
- 促进应激保护代谢物的产生:干旱信号促使细胞产生如脯氨酸和海藻糖等应激保护代谢物。脯氨酸能调节细胞的渗透压,使细胞在缺水环境下保持水分,防止过度失水;海藻糖可以稳定细胞膜和蛋白质的结构,保护细胞免受干旱损伤,帮助细胞维持正常的生理功能。
- 激活抗氧化系统维持氧化还原稳态:干旱会使细胞内产生过多的活性氧物质(如超氧阴离子、过氧化氢等),这些物质会破坏细胞内的生物分子,影响细胞正常功能。为了应对这种情况,干旱信号会触发细胞的抗氧化系统。抗氧化系统中的各种抗氧化物质(如超氧化物歧化酶、谷胱甘肽等)和抗氧化酶协同工作,将活性氧物质转化为无害或危害较小的物质,从而维持细胞内氧化还原反应的平衡状态,即氧化还原稳态,保护细胞免受氧化损伤。
- 利用过氧化物酶防止细胞急性损伤并维持膜完整性:过氧化物酶是抗氧化系统的重要组成部分。在干旱环境下,过氧化物酶被激活,它能够催化过氧化氢等过氧化物的分解反应,将这些具有强氧化性、可能对细胞造成急性损伤的物质转化为水和氧气,从而减轻对细胞的损害。同时,过氧化物酶还能通过参与一些反应来稳定细胞膜的结构,维持细胞膜的完整性,确保细胞内物质不会泄漏,细胞外有害物质不会随意进入细胞,保证细胞正常的生理活动得以进行。
水分胁迫程度以及感知胁迫的植物器官等因素,也会引发特定的信号传导反应,这些反应涉及的信号通路包括但不限于脱落酸、油菜素内酯和乙烯等植物激素通路。当植物遭遇干旱时,干旱的严重程度会影响植物的应对方式。例如,轻度干旱和重度干旱引发的植物反应存在差异。同时,不同的植物器官对干旱胁迫的感知和响应也有所不同。根和叶在感知干旱信号后,激活的信号传导途径和产生的生理反应并不完全一样。这些因素会触发植物体内特定的信号传导,脱落酸(ABA)在植物应对干旱胁迫中起着关键作用,能促进气孔关闭,减少水分散失;油菜素内酯(BRs)参与调节植物的生长发育,在干旱胁迫下,通过与其他激素信号通路相互作用,影响植物对干旱的适应性;乙烯(ETH)则可以调节植物的衰老和防御反应,在干旱条件下,乙烯信号通路被激活,有助于植物调整生理状态以应对干旱。除了这几种激素通路,还有其他信号通路也参与其中,共同调控植物对干旱胁迫的响应 。
对干旱响应基因的过度表达通常会致使植物出现生长缺陷和产量损失,而干旱响应性状的组织特异性或时间特异性表达则有望在不降低产量的前提下,提升植物对干旱的应对能力。尽管水资源的不安全性日益增加,但将包括对基因表达进行调控等多种策略相结合,仍可能实现农业产量的提高。
干旱期间,植物系统维持生理水平的水分平衡的策略:
- 增加从土壤中吸收的根系水分(increasing root water uptake from the soil):干旱时,土壤中的水分含量降低。为了获取足够的水分来维持生命活动,植物的根系会采取一些策略来增强对水分的吸收。例如,根系可能会生长得更深入、更广泛,以扩大与土壤中水分的接触面积。一些植物的根系还会增加根毛的数量或长度,根毛是根系吸收水分和养分的重要结构,根毛的增多可以更有效地从土壤颗粒间的空隙中吸取水分。
- 通过关闭气孔来减少水分流失(reducing water loss by closing stomata):气孔是植物叶片表面的微小孔隙,是植物进行气体交换(如吸收二氧化碳和释放氧气)的通道,但同时也是水分散失的主要途径。在干旱条件下,植物会感知到缺水信号,进而促使气孔关闭。当气孔关闭后,水分从叶片表面向外界环境的蒸发就会大大减少,从而有效地保留了植物体内的水分,维持了水分平衡。然而,气孔关闭也会在一定程度上影响植物的光合作用,因为二氧化碳的吸收也会受到限制。
- 调整组织内的渗透过程(adjusting osmotic processes within tissues):植物细胞内的渗透压对水分的吸收和保持起着关键作用。在干旱时,植物组织会调整细胞内的渗透压,以适应外界环境的变化。植物细胞会在细胞内积累一些溶质,如脯氨酸、糖类等,这些溶质的增加会使细胞内的渗透压升高。根据渗透原理,水分会从低渗透压的区域流向高渗透压的区域,因此细胞内较高的渗透压会促使水分进入细胞,或者防止细胞内的水分流失到外界环境中,从而维持细胞的水分含量和生理功能。
通过以上这些方式,植物系统在干旱期间努力维持着生理水平的水分平衡,以适应干旱环境并尽可能地保证自身的生存和生长。
根系对土壤水分变化的响应是细胞生理调控与系统结构重塑的协同结果:细胞通过渗透调节、信号传导和生长控制维持基本功能,而整个根系通过形态重塑(如主根伸长、侧根 / 根毛分化)和维管优化,在土壤中构建更高效的水分捕获网络。这种跨尺度的适应机制确保植物在多变的水分环境中维持水分平衡,是干旱胁迫下生存与生长的关键策略。
更长、更深且分支角度更小的根系能够有效地从表层干燥但深层保持水分的土壤中捕获水分。
##############################################################################################################
当根系处于水分分布不均的土壤环境中时,会通过水模式化(hydropatterning)过程调整侧根的发生方向,优先向水分含量更高的土壤斑块生长,这一过程由生长素信号通路介导,具体机制如下:
一、水分异质性环境的感知与信号启动
-
局部水势差异的感知
- 根系通过根尖细胞或皮层细胞感知土壤微环境中的水势梯度(如高水分斑块处水势较高),触发细胞膜上的机械敏感离子通道或受体样蛋白(如类受体激酶),激活下游信号(如钙离子波动、活性氧信号)。
- 水分充足区域的根系细胞可能通过渗透调节维持较高的膨压,作为“有利生长区域”的信号被识别。
-
生长素信号的启动
- 水势差异诱导根系局部生长素(IAA)的合成或运输重组。例如,水分丰富的土壤斑块附近,根系中生长素合成基因(如YUCCA家族)表达上调,或生长素从其他区域向该斑块定向运输。
二、生长素介导的侧根原基定向发育
-
生长素的极性运输与局部积累
- 生长素通过极性运输载体(如PIN家族蛋白)在根系中定向运输。在水分高的斑块对应的根区,PIN蛋白的亚细胞定位改变(如PIN3在根尖中柱鞘细胞的重新分布),导致生长素在侧根原基起始位点附近积累。
- 生长素响应报告基因(如DR5:GUS)在水分丰富区域的侧根原基中表达显著增强,表明该区域生长素浓度升高,触发侧根起始。
-
侧根原基的激活与发育
- 高浓度生长素诱导中柱鞘细胞重新进入细胞周期,启动侧根原基的形成(关键基因如ARF7/19介导生长素信号转导,激活LBD家族转录因子,促进细胞分裂)。
- 水分充足的环境中,侧根原基突破皮层的阻力降低(细胞膨压较高、细胞壁松弛酶活性增强),加速侧根的伸出和伸长,形成更多朝向水斑块的侧根。
三、生长素信号与其他通路的协同作用
-
与乙烯、细胞分裂素的交互调控
- 生长素可能抑制乙烯合成(如下调ACS基因表达),解除乙烯对侧根发生的抑制作用,尤其在水分有利区域促进侧根发育。
- 细胞分裂素(CTK)作为拮抗信号,在干旱区域维持较高水平,抑制侧根原基激活;而在水斑块中,CTK信号被生长素下调,允许侧根发生。
-
表观遗传与细胞分化的协调
- 生长素通过组蛋白修饰(如H3K4三甲基化)激活侧根发育相关基因(如SCR、SHR),同时抑制细胞分化相关基因,维持中柱鞘细胞的分裂能力,确保侧根原基在水分有利区域优先形成。
四、生态适应意义
这种由生长素介导的水模式化反应使根系能够高效利用异质性水分资源:在表层干燥但深层或局部存在水斑块的土壤中,侧根集中在湿润区域发育,扩大水分吸收表面积,同时减少在干旱区域的无效生长,优化碳资源分配。例如,拟南芥在水分斑块化土壤中,侧根密度在高水区域可增加50%-80%,显著提升水分捕获效率。
总结
根系的水模式化是生长素信号主导的适应性策略:通过感知水分梯度,重组生长素运输与分布,定向激活侧根原基发育,使侧根优先向水分丰富的土壤斑块延伸。这一过程整合了局部水分信号、激素调控网络和细胞分裂分化事件,体现了植物根系在异质环境中“趋利避害”的精准调控机制。
#############################################################################################################
脱落酸(ABA)作为一种植物激素,能够调控植物对脱水胁迫的响应并优化水分利用。脱水信号会刺激植物不同器官局部合成 ABA,但叶片叶肉细胞中 ABA 的合成效率显著高于根组织。积累的 ABA 随后激活下游信号组件。在胁迫过程中,ABA 通过介导与其他信号通路的交叉对话来执行其功能(图 3)。许多现有的提高水分利用效率和抗旱性的策略均涉及 ABA 信号通路。
油菜素甾醇类激素也通过与 ABA 响应通路相关的信号组件调控干旱反应(图 3)。油菜素甾醇信号负调控因子 —— 油菜素甾醇不敏感蛋白 2(BIN2)可被 ABA 不敏感蛋白 1(ABI1)和 ABI2 去磷酸化。ABA 通过抑制 ABI1 和 ABI2 的活性来激活 BIN2。活化的 BIN2 对 SnRK2 蛋白进行磷酸化修饰,进而激活下游通路。ABA 信号还可在下游转录因子水平与油菜素甾醇通路汇聚(图 3)。油菜素甾醇信号中的 BRI1-EMS 抑制因子 1(BES1)会抑制 ABA 诱导的干旱相关转录因子 —— 脱水响应蛋白 26(RD26)的表达。反之,RD26 通过调控 BES1 介导的转录并抑制油菜素甾醇调控的生长,与油菜素甾醇形成双向拮抗关系。
WRKY46 是植物整合油菜素甾醇与脱落酸信号的关键转录因子,通过与 BES1 互作促进生长并抑制干旱响应,而干旱时被 BIN2 介导的磷酸化降解所抑制,从而释放抗逆程序。这一机制体现了植物在资源有限条件下的生存策略优化,为抗旱作物育种提供了靶向 WRKY 家族转录因子的新思路。
改善根系的水分获取能力也能提高植物在干旱条件下的生长表现。在拟南芥中,通过全基因组关联分析鉴定出了调控根系系统深度的生长素通路调控因子 —— 外囊复合体亚基 EXO70 家族蛋白 A3(EXO70A3)(40)。作为胞外分泌系统的组成部分,EXO70A3 在根尖表达,通过影响根冠柱细胞中生长素外排载体 PIN-FORMED 4(PIN4)的稳态来调控局部生长素运输(图 3)。EXO70A3 的自然变异与季节性降水相关,并在不同降雨模式下赋予植物不同的适应性根系结构构型。在高温灌溉地区,更深的根系结构被证明更有利于干旱适应。在水稻中,生长素诱导基因DEEPER ROOTING1通过促进形成更垂直且更深的根系结构来增强抗旱性。尽管生长素在胁迫条件下调节根系结构,但根系的向水性响应功能相对独立于生长素,并涉及根伸长区的脱落酸(ABA)信号传导。在根伸长区皮层细胞中,脱落酸(ABA)诱导的 MIZU-KUSSEI1(MIZ1)与 SnRK2.2(SNF1 相关激酶 2)的协同作用可解读土壤环境中的水势梯度。
在干旱条件下,根系感知土壤中的水分短缺,植物地上部分通过关闭叶片气孔作出响应,这表明存在一种系统性的通讯机制。干旱时,根系产生 CLE25 肽,该肽通过维管系统运输至植物叶片,通过激活生物合成酶 NCED3 驱动脱落酸(ABA)的合成。ABA 合成的激增导致气孔关闭并改善水分平衡,从而促进植物在干旱中的存活(图 3)。对拟南芥中小分子肽信号传导的这一发现,可能有助于在作物中鉴定类似的机制,以实现从根系到地上部的胁迫信号传递。
重要图表
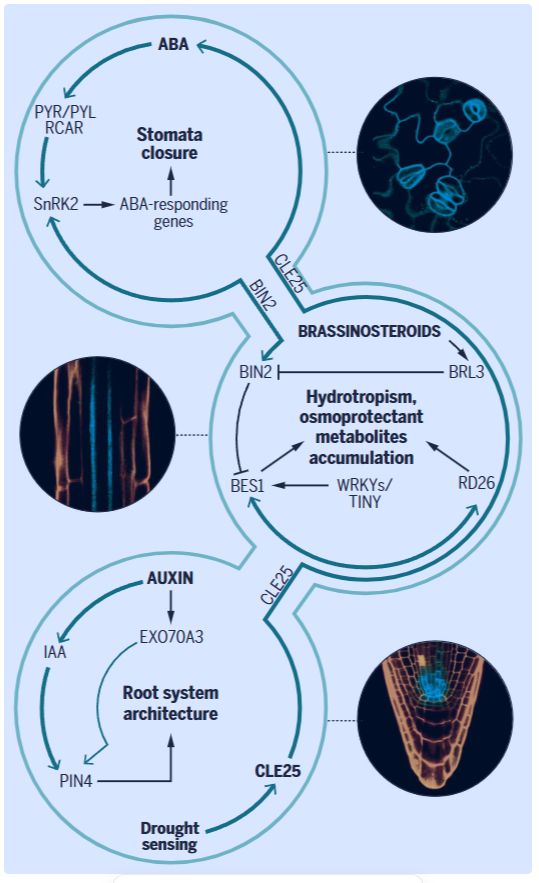
图 3. 支撑干旱响应的激素信号事件。调控干旱适应性的激素信号模块示意图。植物通过叶片、维管组织和根系等器官对抗脱水。脱落酸(ABA)通过 SnRK2 激酶激活多种基因,促使气孔关闭并改善水分平衡。当根系感知干旱时,CLE25 肽通过维管系统运输至叶片,在局部调控 ABA 的生物合成及气孔关闭。油菜素甾醇类激素也在调节植物干旱响应中发挥作用:油菜素甾醇通路通过下游组分 BIN2 激活 SnRK2,从而与 ABA 通路汇聚,反之亦然。独立于 ABA 之外,油菜素甾醇受体(BRI1、BRL1 和 BRL3)可调控根系的向水性响应 —— 维管组织中的 BRL3 受体通过促进根系组织中渗透保护代谢物(如脯氨酸、海藻糖等)的积累,协调干旱胁迫下植物的生长与存活。此外,通过 EXO70A3 和 PIN4 介导的非经典生长素响应可调节根系结构模式与生长深度,从而增强从土壤中吸收水分的能力,进而提升耐旱性。