异质脑:自闭症谱系障碍患者自发连接模式畸变
来自以色列魏茨曼科学研究学院的Avital Hahamy等人在Natureneuroscience上发表文章,发现自闭症谱系障碍(Autism spectrum disorders,ASD)患者大脑静息态时功能连接模式异常。以往研究发现的ASD伴随静息态功能连接减弱的断言最近被增加的功能连接挑战,针对这些矛盾的发现,作者检验了来自几个静息态数据中心的高功能ASD成年患者和匹配的正常对照成年人半球间和半球内的功能连接。结果在多组ASD中同时发现了增强和减弱的功能连接。作者提出这种异质性源于之前未被识别的ASD特性:功能连接模式相对于标准模板的异质畸变。并且个体等位半球间连接模式畸变幅度与ASD行为症状相关。作者提出功能连接组织的改变是高功能ASD的核心特性,并且可能是以往矛盾发现的原因。
背景
静息态时,典型成年人脑呈现活动模式和脑网络的相关性。功能磁共振(fMRI)和电生理记录均发现典型成年人脑半球内和半球间自发功能连接。但这种功能连接在皮层上并不是均匀的,例如,初级感觉运动区显示高水平的连接,而更高级的联合区连接程度稍弱。类似的,每个网络内不同区域间功能连接也不同。
自闭症谱系障碍(Autism spectrum disorders,ASD)是基于社交语言能力损伤以及大量重复行为诊断的神经发育疾病。大量研究表明ASD大脑非典型性功能连接模式。非典型性包括多种功能连接的减弱,尤其是半球间连接。但是,这种“连接不足”被最近报道的“过连接”研究挑战。
该研究用大样本fMRI静息态扫描来寻找可能解决这些矛盾的潜在原理。结果发现新的鲁棒的ASD连接异常:功能连接模式的拓扑特性而非全局强度。特别的,正常对照中功能连接典型模式在孤独症患者中显示出重要的且个体不同(异质)的畸变。这种异质特性导致ASD个体间连接模式的高度差异,包括半球内和半球间连接模式。半球间等位连接个体畸变的幅度与ASD患者的行为测量相关。
结果
半球间等位连接
作者首先分析了等位半球间连接,因为它们提供了相对简单却鲁棒的脑连接。并且,不同于功能连接的矛盾发现,这类连接被一致报道在ASD中降低。共检验了5个中心ASD组相比于正常对照组体素水平的差异,计算了每个中心ASD组和正常对照组全局半球间连接的比例。结果发现所有比值均稍小于1,说明ASD组微弱却一致的全局半球间连接的减弱。但这种效应并没有通过多重比较校正。
为了估计ASD样本等位半球间连接不足是否在皮层拓扑均匀 ,作者检查群组水平半球间连接图谱。与之前报道一致,作者在正常组和ASD组均发现了遍布皮层的半球间连接强度局部差异模式。最高的半球间连接位于初级感觉运动皮层(中央前回,中央后回),紧接着是其他感觉皮层(枕叶,横回),显示相对低的半球间连接的区域是异模态联合区域,如额叶和颞叶(颞中回,颞下回)。但是,即使定性评估都显示ASD组和正常组半球间连接的显著差异。
为了定量评估ASD组和正常组半球间连接差异,对于每个中心及合并样本分别做了组间T 检验,结果ASD组既显示了连接增加的区域又显示了连接减弱的区域(图1.c)。采用荟萃分析来估计不同中心连接的一致性,结果仍显示了连接增加和连接减弱的区域。
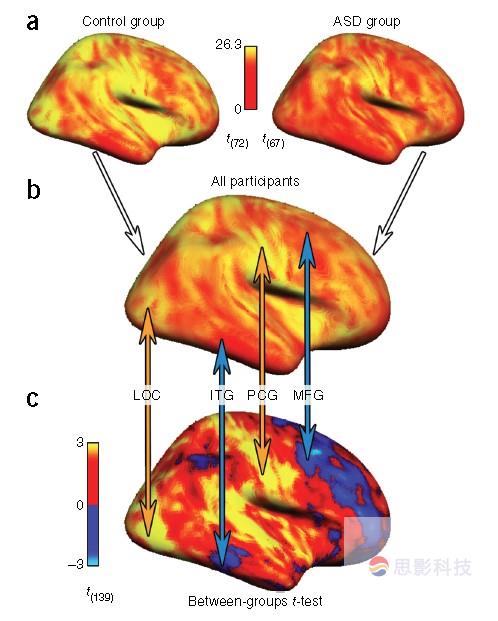
图1.合并的等位半球组图谱和组间差异图谱。(a)对照组(n = 73)和ASD组(n =68)等位半球图谱,整合了所有数据集。(b)所有数据集对照和ASD患者平均等位半球图。(c)合并的组间t检验图谱(对照 >ASD)。箭头显示了组差异的方向和等位半球连接的局部变化。ASD组在高半球连接区域显示降低的半球间连接(橙色箭头),在低半球连接区域显示增强的半球间连接(蓝色箭头)。等位半球连接图谱以中线对称,所以只显示了右半球。LOC,外侧枕叶皮层;ITG,颞下回;PCG,中央后回;MFG,额中回。
组差异的空间结构
为了寻找连接不足和过连接效应图谱潜在的原理,作者在合并样本集用典型半球间连接图比较了群组间t图。因为每个实验组的均值和t检验得到的均值间的差异在数学上是独立的,作者在所有141个被试比较了平均半球间连接图(图1.b,c)。比较发现,组差异的方向(control > ASD 或者 ASD >control)由半球间连接的典型数值决定。具体的说,半球间连接高的区域(如,感觉运动皮层,枕叶皮层)在ASD中相比正常对照连接降低。而在半球间连接低的区域(如,额叶,颞叶)在ASD中有增加的功能连接。作者将这种现象称作“向均数回归”效应。
在特定数据集中也发现了这种“向均数回归”效应。为了避免循环,在比较组间差别图和正常组图时排除了计算组间差异图时用到的正常被试。结果再次显示高半球间连接的区域在ASD病人中显示了低水平的连接,同样在低半球间连接的区域出现了相反的结果。
“向均数回归”效应的直观预测是在ASD组图谱中极端体素值(包括高低半球连接)相比于正常组图谱中体素值会衰弱,这将导致体素连接值的方差下降。在所有组中都发现了这种效应,确认了“向均数回归”效应的出现(P =0.01)。
“向均数回归”效应起源
群组水平的“向均数回归”效应可能来源于个体水平的两个不同现象。首先,在组水平效应可能来源于ASD组个体的“向均数回归”。为了验证,将应用于组图谱的定量测量用于单个被试图谱并且测量单个脑连接值半球间连接的方差。但是,单个被试方差的组平均的比例在不同数据集并未显示组间一致效应,并且在大多数比例表现了相反的趋势(ASD组高方差),说明在组水平“向均数回归”效应并非来源于单个个体水平“向均数回归”。
另一个可能的解释是“向均数回归”组效应来源于ASD患者组间半球间连接有更高的个体拓扑畸变。图2描述了这种个体畸变为什么会导致组水平“向均数回归”。半球间连接值的平均个体方差在正常对照和ASD患者间无差别。相关,被试间高连接幅度和低连接幅度的未对准导致了高幅度和低幅度的混合,导致组水平“向均数回归”效应。

图2. “向均数回归”效应空间畸变来源,红-黄表示高的等位半球连接,蓝白表示低的等位半球连接。组图谱由单个被试图谱叠加生成。正常组低的空间变化导致组水平高低半球间连接的不同区域(左)。但是,在ASD患者中,异质空间畸变导致组图谱中“向均数回归”效应(右):半球间连接高的区域显示低的连接水平,相反的情况成立。柱状图显示了每行两个图谱的定量比较。每个柱代表假设高半球连接的体素的值(H)和假设低半球连接的体素的值(L)。垂直线展示了每个图值的散布。“向均值回归”效应在组水平非常明显因为ASD组值的低离差。这个效应并非来自于单个被试水平,因为在ASD组单个被试内值的离差于对照组无差别。
如果这种可能性存在,不同的ASD个体需要表现半球间连接的异质模式:每个个体显示一个典型模式的不同畸变。为了评估这种可能性,在每个实验组计算了被试半球间连接的空间相关性,得到的相似性矩阵一致表明正常对照组相比ASD组高的连接值(图3.a)。ASD组个体全局连接模式相似性与正常组比例均一致得小于1,证明在ASD组更多样的半球连接模式。因此,作者明确的指出ASD患者个体间更大范围的异质连接模式是导致群组半球间连接模式的“向均数回归”的潜在现象。
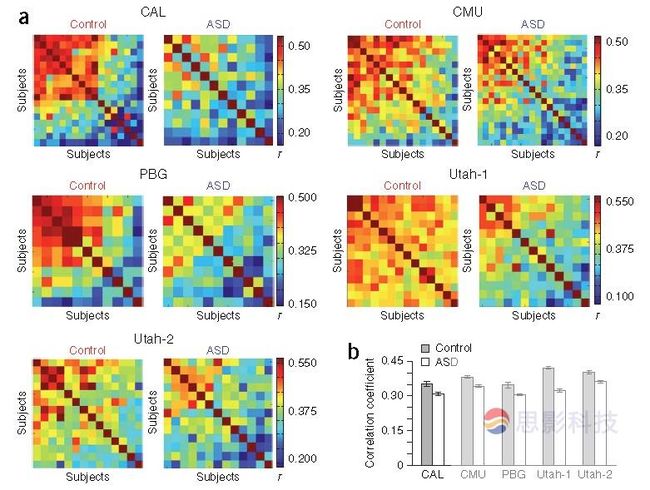
图3. ASD组等位半球间连接的异质分布。(a)分别对于每个中心,对正常组(left)和ASD组(右)呈现相似性(空间相关)矩阵。每行每列代表一个被试和该试验组其他所有被试等位半球连接图谱的相似性。每个矩阵内被试按照平均相似性值降序排列。注意正常对照组相比ASD组个体间高相似性。(b)a图中数据集对照组和ASD组每个矩阵平均被试间相似性值。Error bar代表标准误。所有数据集正常对照组平均被试间相似性均大于ASD组,说明ASD样本相比于正常样本等位半球连接的异质畸变( χ 2 (10)=31.95,P = 0.01校正,Fisher‘s 方法)。
功能连接模式的替代选择
作者已经说明在ASD患者中,半球间连接的空间模式相似性更低。但是,很可能在不同数据集在ASD患者中出现相似的空间畸变。这个假设需要ASD患者有标准的ASD模式(包含系统模式畸变)而非一个标准的正常模式(没有这些畸变)。然而,在目前数据中,作者并未发现这种效应(t(67) =-0.81,P = 0.42)。
作者关注ASD患者的异质模式,还有一种可能是在ASD群体存在几种不同模式,这可能会使ASD个体构成子组。这些模式在子组内应该相似但在子组间不同。为了验证,作者将ASD和正常被试等位半球间连接分别用K-means 聚类并且用gap
statistics检验得到的团簇。为了避免因为不同中心和采集序列导致的伪团簇,将聚类分别应用到每个数据集。结果显示不同中心正常组和ASD组的最优团簇数目都是1。因此,并未显示ASD组相比于正常组在形成团簇上的差异。
最后,ASD组的个体差别也可能来自于解剖差别。比如,ASD患者可能显示更高的不对称性。但是,结构不对称上的组差异非常小,在不同中心间也不一致。因此,不太可能是这个原因导致了功能测量的偏差。
与ASD诊断测试的关系
作者进一步假设ASD患者大脑连接模式畸变的水平与行为症状相关。为了给每个ASD患者绘制半球间连接的拓扑畸变,对于每个个体,在体素水平减去对照组半球连接值。得到的单个被试图谱表示每个体素相对于典型半球间连接模式的偏差程度。图4a代表ASD诊断观察量表(Autism
Diagnostic Observation Schedule,ADOS)极值的两个个体(最低和最高ADOS总分数)。即使是定性来看,ASD症状最严重的个体表现出更大的离差,相比于严重程度稍弱的典型的半球间连接模式既有正值也有负值。换句话说,症状最严重的患者相比对照模式偏差的方差更大。
因此作者定义了一个变形指标描述对于每个个体脑半球间连接水平差别的方差。发现了变形指标和ADOS测量显著的正相关。但在Bonferroni校正后,仅有和ADOS总分数(r = 0.32, P =
0.045,校正后, n = 66)及交流分数(r =0.33,P = 0.04,校正后,n =66)的相关性存在(ADOS社交P =0.13,刻板行为P =0.5,校正后)。作者也计算了对于对照被试的畸变水平,同预期一致,正常组的畸变水平低于ASD组(ASD/对照的畸变比例1.13, P = 0.01)。
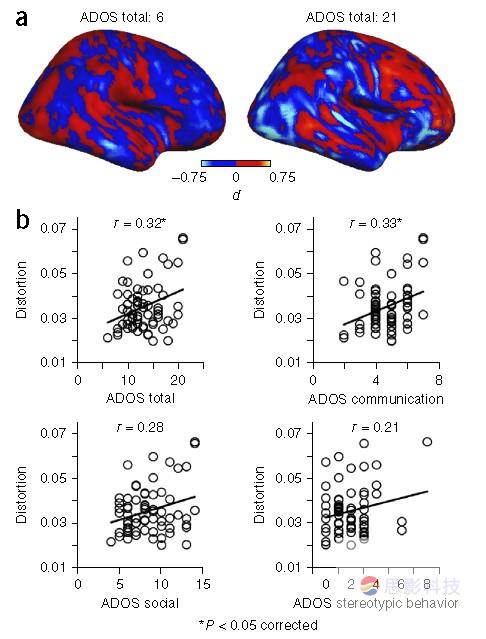
图4.个体模式畸变定量化与ASD症状相关性。(a)ASD单个被试值减去平均等位连接对照值,这里展示两个最低(左)和最高(右)ASD总分数的两个被试。注意低ASD分数的被试等位半球连接差别值最小,表明低水平畸变。相反的情况存在于高ASD分数的个体。(b)单个被试ASD指标与单个被试ASD症状的相关性。每张图都有相关系数。尽管所有的系数在多重比较校正前都显著,但只有ASD总分数和ASD交流能力(上面一行)通过校正。
异位半球间和半球内模式
异质连接畸变可能并不止独特于等位半球连接而是反应更一般的脑现象。作者比较了异位半球模式,和正常组及ASD组半球内连接模式。相比正常组,ASD组异位半球模式显示了被试间空间相似性的一致减弱。在左右两个半球中,半球内连接模式也发现了同样的现象。
进而对于异位半球模式也计算了单个被试畸变指标与临床ASD测量、年龄和IQ分数的相关性。在半球间连接和右半球内连接中均发现了显著的正相关,但均未能通过多重比较校正。
如果连接不足是ASD脑的全局特征,在ASD组所有的脑连接相比正常组均应显示低幅值。但是,不像半球间连接的情况,在异位半球模式和左右两个半球内模式上均未显示一致的全局连接不足效应。
讨论
ASD空间畸变连接模式
该研究显示了高功能自闭症的成年人脑异常的新特性,作者发现ASD患者静息态功能连接的拓扑模式相对于典型对照显著畸变,并且这种功能连接模式畸变在ASD个体异质程度高于正常对照被试。
对于拓扑变形现象最直接的证据是ASD个体相比于正常对照低个体间相似性。在不同的连接类型中,均一致得发现了这种效应。在每个半球的功能连接模式和半球间的功能连接模式。
增加的空间多样性导致了“向均数回归”效应来源于ASD组等位半球间连接方差下降。值得注意的是,组水平“向均数回归”可能来源于ASD个体两个半球等位体素的未对齐。但是,这种未对齐必然导致个体水平极端半球间连接值的衰减(“向均数回归”),实际并不是这样。
ASD个体增加的个体变异可能来源于ASD不同亚型。但是在等位连接模式,并未发现这种亚型。可能的连接模式非常多,很可能需要大的数据集来发现这种聚类。
与行为测量的联系
以前的研究将ASD功能连接改变当作分类性状(区分ASD和非ASD个体),但其他的神经生理参数被当作维特征(将ASD放在正常人连续谱的末端)。该研究的结果兼容这两个方面,因为异质的功能模式可以区分ASD个体和非ASD个体,但是异质的程度与ASD在ADOS测量上的严重程度对应。需要进一步的神经和行为比较来确定ASD中报道的功能异质是一般群体功能模式在维度上分布的的极值。
不同于在成年人中反映ASD症状的ADOS分数,自闭症访谈量表(ADI)分数并未与异质畸变分数显著相关。但是,因为仅有将近一半的被试有ADI分数,不太可能确定汇报的效应是否有发育起源,或者与代偿机制相关。一并考虑异质畸变和年龄的相关性,需要在儿童ASD上进一步研究来处理本研究发现的发育方面。
行为测量和半球异位及半球内空间畸变的相关性仅有不明显的趋势,尽管和半球同位的发现方向一致。这显示半球同位连接模式是一个更敏感的标记,可能要归因于相比于其他脑连接信噪比高。作者也注意到异质模式和行为症状相关的显著性比其他效应弱。众所周知,ASD的行为测量很难定量,这些相关中变化的来源仍需要阐明。
畸变程度和IQ并没有发现显著的相关。这说明,尽管在实验组间发现了显著的IQ差别,但IQ差别并不是组间畸变差别的来源。
与之前功能连接发现的联系
该研究并没有发现脑连接的不同类型显著的全局连接不足效应。但是,我们的结果显示了在ASD组相比正常组增加和降低的功能连接均可能导致个体水平的连接模式的空间畸变。在等位半球连接时,个体间平均畸变模式导致高和低连接模式明显“稀释”,在ASD组反映出“向均数回归”效应。因此要小心解释正常个体和ASD个体局部组差异,因为这可能跟ASD异常连接模式相关。
多中心数据使用
在所有ASD分析样本中发现的连接异质空间畸变证明了这种效应的鲁棒性,尤其是考虑了数据集间的变化。尽管我们并没有控制实验相关的差别源(例如,不同的实验序列),连接模式的空间畸变都足够断言超过任何方法上或人口统计上的差异。
另外,作者在不同中心大样本个体上描述了功能连接特性使得作者可以控制其他可能解释之前研究不一致发现的变异来源。首先,作者通过将研究对象锁定为高功能成年ASD患者限制了ASD被试人口统计和临床特性的变化。其次,不同中心的数据都用了相同的分析方法。最后,通过采用方法降低头动对功能数据的影响仔细地控制了非神经元对于fMRI信号的贡献。
异质功能模式的可能来源
什么样的原因可能引发ASD自发连接模式特异性呢?当考虑等位半球畸变,一个直接的解释是在ASD中半球结构偏侧化改变。另一个可能性是ASD个体具有非典型脑。在这种情况下,向标准空间的变换可能会引起改变的解剖。但是,并没有发现在畸变水平转换到标准空间引起的组差别,挑战了这个选择。
进一步的可能性是组差别与任务相关的神经活动异质差别相关。在成年人和青少年高功能ASD中,一些研究与这种可能性兼容。特别的,在简单的运动任务、更复杂的视觉运动学习、面孔处理、自然感觉刺激中半球内和半球间的异质激活有被发现。然而这些研究大多在相对下的样本中检查在设计好的实验任务下特定脑区的活动。该研究提供了一个可以解释之前研究的更一般的框架。换句话说,在大样本数据中,所有脑区和连接类型中出现的异质功能连接模式(即使在没有外显任务下也出现),提出一个新的ASD功能特异。
进一步,没有外显的任务下出现的异质性是非常有意思的,因为自发出现的功能连接模式被日常出现的个体行为塑造。如之前假设,与外界环境的交互驱使的惯常行为可以被编码为突触效能,并且在没有外显行为时出现。ASD个体异质连接模式同样可能来源于ASD个体与外界交互的改变。在社交和物理环境因素及他们引起的脑激活在典型的个体由不同个体共享假设下,环境-神经元的交互性会调整网络组织,导致趋同标准脑。相反,ASD个体与调整后的组织行为分离,加上他们证据充分地对于外界环境不可靠地和过可塑性响应,会导致ASD个体大脑异常和异质的连接模式。值得进一步研究的一个有趣的问题是这种连接异质是否会出现在显示非正常脑-环境交互的其他临床群体。
参考文献:Hahamy, A., M. Behrmann, and R. Malach, The idiosyncratic brain: distortion of spontaneous connectivity patterns in autism spectrum disorder.Nature neuroscience, 2015.18(2): p. 302-309.
微信搜索“思影科技”公众号,第一时间获取脑影像资讯