- 【Linux系统学习】1.初识Linux
A 北枝
Linuxlinux学习运维
初识Linux操作系统概述初识Linux虚拟机介绍VMwareWorkStation安装1.操作系统概述了解操作系统的作用了解常见的操作系统1.1硬件和软件计算机由哪两个主要部分组成?硬件:计算机系统中由电子,机械和光电元件等组成的各种物理装置的总称。软件:是用户和计算机硬件之间的接口和桥梁,用户通过软件与计算机进行交流。而操作系统,就是软件的一类。一个完整的计算机:1.2操作系统操作系统是什么?
- 基于c++图形化界面的clock页面置换算法
LEMFOooO
一、课程设计目的《操作系统》是一门重要的专业基础课,是涉及较多硬件知识的计算机系统软件课程。在计算机软硬件课程的设置上,它起着承上启下的作用。操作系统对计算机系统资源实施管理,是所有其他软件与计算机硬件的唯一接口,所有用户在使用计算机时都要得到操作系统提供的服务。操作系统课程设计的主要任务是研究计算机操作系统的基本原理和算法,掌握操作系统的进程管理、存储管理、文件管理和设备管理的基本原理与主要算法
- 第九届“中国软件杯”大学生软件设计大赛总决赛落幕
趣味科技v
人工智能大数据物联网机器学习交互设计
11月3-5日,第九届“中国软件杯”大学生软件设计大赛(以下简称大赛)总决赛在江苏常州圆满举办。经过九年积累,大赛已经吸引包括全部985、211高校在内的千余所院校、数万支团队、二十余万名软件与计算机及相关专业学生参赛,赛题紧紧围绕企业际需求,已帮助国内软件骨干企业解决共性技术难题百余道。大赛已经成为提升学生实践创新、促进软件校企对接、推动产教融合共育人才的重要平台。总决赛期间,包括决赛评审、产教
- 整理1
小赵不在
操作系统定义:操作系统是一个管理和控制计算机硬件与软件资源的计算机程序。位置:操作系统工作在计算机硬件之上,软件工作在操作系统之上。(中间位置)作用:用户和软件与计算机硬件之间传递信息的系统软件。分类:1、桌面操作系统Windows(用户群体大)macOS(适用于开发人员)Linux(应用软件少)2、服务器操作系统Linux(安全、稳定、免费、占有率高)WindowsServer(付费、占有率低)
- 软件开发流程
阿瞒有我良计15
测试开发运维
目录软件软件开发流程的演变瀑布模型敏捷模型XPSCRUMDevOps1.软件与计算机系统操作有关的计算机程序、可能有的文件、文档及数据。软件可以分为两种主要类型:独立软件:独立软件是一种完整的应用程序,可以直接在计算机或移动设备上安装和运行。它们通常有一个用户界面,用户可以通过界面与软件交互并执行特定的功能。例如,办公软件套件、图形图像编辑器、游戏等。嵌入式软件:嵌入式软件是嵌入到硬件设备中的软件
- 学习《数据结构》要爬的第一步梯子
宝藏女孩的成长日记
数据结构与算法数据结构算法
小王开始专业课的复习一、数据结构是什么二、数据结构中基本概念和术语2.1数据结构的概念2.2按照不同功能划分逻辑结构2.3存储结构中重点顺序存储举例链式存储举例三、抽象数据类型的表示与实现3.1数据类型3.2抽象数据类型(ADTs:AbstractDataTypes)3.3算法分析(重点)1.时间复杂度2.空间复杂度复习数据结构正式开始!一、数据结构是什么数据结构是介于数学、计算机软件与计算机硬件
- 划重点!“芯片、软件与计算”(芯片类)重大专项项目可申报了!
深科信
省直有关部门、各地级以上市科技局(委)、各有关单位:为全面贯彻落实党的十九大和习近平总书记关于加强关键核心技术攻关的系列重要讲话精神,按照省委省政府关于科技创新的相关部署,根据《广东省重点领域研发计划实施方案》,现启动2020年度广东省重点领域研发计划“芯片、软件与计算”(芯片类)重大专项项目申报工作(申报指南见附件1)。有关事项通知如下:一、申报要求(一)项目申报单位(包括企业、科研院所、高校、
- 学习第一天
辰王书
day1什么是计算机?1.构成:软件与硬件(1)软件与计算机系统操作有关的计算机程序、规程、规则,以及可能有的文件、文档及数据。分为系统软件(系统软件为计算机使用提供最基本的功能,可分为操作系统和系统软件,其中操作系统是最基本的软件。)与应用软件(系统软件并不针对某一特定应用领域,而应用软件则相反,不同的应用软件根据用户和所服务的领域提供不同的功能。)操作系统:操作系统是一管理计算机硬件与软件资源
- C++计算机软件系统
Joseph_tony
c++c++
计算机软件是指计算机系统中的程序及其文档,也是用户与硬件之间的接口,用户主要通过软件与计算机进行交流,软件是计算机的灵魂。没有安装软件的计算机称为“裸机”,无法完成任何工作。一般软件分为系统软件和应用软件两大类。硬件位软件提供运行平台。软件和硬件相互关联,两者之间可以相互转化、互为补充。计算机的软件分成系统软件和应用软件两大类。系统软件系统软件是指控制和协调计算机及外部设备,支持应用软件开发和运行
- OpenBLAS简介及在Windows7 VS2013上源码的编译过程
fengbingchun
CaffeEigen/OpenBLAS
OpenBLAS(OpenBasicLinearAlgebraSubprograms)是开源的基本线性代数子程序库,是一个优化的高性能多核BLAS库,主要包括矩阵与矩阵、矩阵与向量、向量与向量等操作。它的License是BSD-3-Clause,可以商用,目前最新的发布版本是0.2.19。它的源码放在了GitHub上,由张先轶老师等持续维护。OpenBLAS是由中科院软件所并行软件与计算科学实验室
- 厦大计算机研究生
loverszhaokai
考研
140软件学院35本单位35名招生计划中,软件工程(专业学位)招生计划为15名,学术型专业计算机软件与理论招生计划为20名。081202计算机软件与理论①101思想政治理论②201英语一③301数学一④408计算机学科专业基础综合06数据挖掘技术及其应用07智能软件与计算理论08模式识别与图像处理09数据库理论及其应用技术10软件工程与面向对象设计11数字媒体技术085212软件工程(专业学位)①
- 课堂学习内容
zhou_mumu
软件的定义软件是用户与硬件之间的接口,用户主要通过软件与计算机交流软件的分类按功能分类1.系统软件(1)操作系统(2)支撑软件2.应用软件按软件的规模分类微型,小型、中型、大型、甚大型和极大型6种按工作方式分类(1)实时处理软件(2)分时软件(3)交互式软件(4)批处理软件软件架构1.软件架构的定义软件架构是有关软件整体结构与组件的抽象模式,用于指导大型软件系统各个方面的设计。软件架构是一个系统的
- 专业导论-课后总结-2019.12.05-V1.0
不想写博客ト
课后总结
3.1、软件的定义软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件=程序+数据+文档程序:按照事先设计的功能和性能要求执行的指令序列。数据:使程序能正常运行的参数及程序运行过程中所处理的对象。文档:与程序开发,维护和使用有关的资料总称。3.2、软件的分类3.2.1、按照功能分类3.2.1.1、系统软件(1)操作系统工作职责:决定系统资源供需的优先次序,管理与配置内存,控制输入
- 第十四周课后总结
weixin_45889959
导论课后总结
第十四周课后总结3.1软件的定义用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件是计算机系统中的非硬件成分,它是包括程序、数据及其相关文档的集合。软件是计算机系统设计的重要依据。3.2软件的分类3.2.1按软件的功能分类1.系统软件(1)操作系统:管理计算机硬件与软件资源的程序,同时也是计算机系统的内核与基石,提供了用户与系统交互的操作接口(2)支撑系统:包括一系列基本工具,以及数
- 计算机导论——第四次课后总结
weixin_45816647
第三章走进软件3.1软件的定义软件是用户与硬件之间的接口,用户主要通过软件与计算机交流。从传统意义上讲,软件是计算机系统中的非硬件成分,是程序、数据、文档的集合3.2软件的分类3.2.1按功能分类可分为系统软件和应用软件1.系统软件操作系统(DOS、Linux、Windows等)、支撑软件(WebSphere、studio.net等)2。应用软件办公、文字处理、绘图、娱乐等3.2.2按规模分类微型
- 第四次课后总结
酷酷的琦
1.软件的定义用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件又是计算机系统中的非硬件成分,它是包括程序、数据及其相关文档的集合。软件还是计算机系统设计的重要依据。2.软件的分类①按功能分类(1)系统软件系统软件由两部分组成,一是操作系统即管理计算机硬件与软件资源的程序,同时也是计算机系统的内核与基石,提供了用户与系统交互的操作接口。二是支撑系统即包括一系列基本工具,以及数据库管理
- 计算机导论第四节课知识总结
传说试驾
计算机导论
计算机导论第四节课知识总结本文仅为应学校要求对上课内容知识点的进行总结,基本上自己总结,概念不全见谅3.1软件的定义软件是用户和软件之间的接口,用户主要是通过软件与计算机进行交流的。3.2软件的分类3.2.1软件按功能分类软件按功能分可以分为系统软件与应用软件1.系统软件系统软件与计算机硬件紧密配合在一起,使计算机系统中的各个部件、相关的软件和数据协调、高效的工作。(1)操作系统:操作系统是管理计
- 计算机导论课后总结
qq_45883825
走进软件1.软件的定义软件是用户和硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件是计算机系统设计的重要依据。2.软件的分类软件分类主要有两种,分别是按软件的功能分类、按软件的规模分类和按软件的工作方式分类;按照软件的功能主要分为系统软件和应用软件。按照软件的规模分类主要是参考所需要的人力、时间、以及完成的源程序代码行数分为6类;按照软件的工作方式分为实时处理软件、分时软件、交互软件和批
- 计算机导论(12.05)课后总结
TianYuZou
计算机导论
3.1软件的定义(1)软件是用户于硬件之间的接口,用户主要是通过软件与计算机进行交流的。(2)国标中对软件的定义为:与计算机系统操作有关的计算机程序、规程、规则,以及可能有的文件、文档及数据。3.2软件的分类3.2.1按软件的功能分类(1)系统软件系统软件为计算机使用提供最基本的功能,可分为操作系统和系统软件,其中操作系统是最基本的软件。1.操作系统操作系统是一管理计算机硬件与软件资源的程序,同时
- 计算机导论第五节课程总结
qq_35008089
总结笔记
软件的定义软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件:与计算机系统操作有关的计算机程序、规程、规则,以及可能有的文件、文档及数据。1.运行时,能够提供所要求功能和性能的指令或计算机程序集合。2.程序能够满意地处理信息的数据结构。3.描述程序功能需求以及程序如何操作和使用所要求的文档。以开发语言作为描述语言,可以认为:软件=程序+数据+文档软件的分类按功能分类1.系统软件
- 计算机导论课后总结-14周-计算机m1804方振杰
caixukun2000
课后总结
软件的内涵软件是用户与硬件之间的接口界面。用户主要是通过软件与计算机进行交流。软件是计算机系统设计的重要依据。为了方便用户,为了使计算机系统具有较高的总体效用,在设计计算机系统时,必须通盘考虑软件与硬件的结合,以及用户的要求和软件的要求。软件的正确含义应该是:运行时,能够提供所要求功能和性能的指令或计算机程序集合程序能够满意地处理信息的数据结构描述程序功能需求以及程序如何操作和使用所要求的文档操作
- 计算机科学导论课后总结_4
Mogana_Lee
计算机科学导论
计算机科学导论课后总结_4一、走进软件#1、软件软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。1、软件分类:按系统的功能分类:系统软件、应用软件。按软件的规模分类按软件的工作方式分类:实时处理软件、分时软件、交互式软件、批处理软件。2、软件架构:有关软件整体结构与组件的抽象模式,用于指导大型软件的各个方面的设计。#2、操作系统1、操作系统定义:系统观点——计算机资源管理在计算机
- 计算机导论总结(四)
LiSeMn
一、走进软件(1)软件的定义:软件是用户与硬件之间的接口,用户主要通过软件与计算机交流。从传统意义上讲,软件就是计算机系统中的非硬件成分,是包括程序、数据及相关文档的集合。(2)分类按功能分:1.系统软件系统软件为计算机使用提供最基本的功能,可分为操作系统和系统软件,其中操作系统是所有软件运行的基础。2.支撑软件3.应用软件应用软件是为了满足某一领域、某类问题的应用需求而提供的软件。按规模分:按照
- 专业导论04
C-Bob
专业导论
专业导论-课后总结-2019.12.083.1、软件的定义软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。软件=程序+数据+文档3.2.系统软件(1)操作系统工作职责:决定系统资源供需的优先次序,管理与配置内存,控制输入输出设备,操作网络与管理文件系统等。常见操作系统:DOS,Linux,MacOS,OS/2,UNIX,Windows应用软件:是为了,满足某一领域,某类问题的应用
- 计算机导论第三章
Agony129
走进软件软件的定义软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。从传统意义上来讲,软件是计算机系统中的非硬件成分,它是包括程序、数据及相关文档的集合。软件的分类按软件功能分类:按软件的功能可以将软件分为系统软件和应用软件。1.系统软件系统软件与计算机硬件紧密配合在一起,是计算机系统中的各个部件、相关的软件和数据协调、高效的工作。系统软件使得计算机用户和其他软件可以将计算机当做一
- 软件的内涵、操作系统、软工定义、程序设计语言的演变
wufixiaoxuxu
软件的内涵、操作系统、软工定义、程序设计语言的演变软件的内涵软件是用户与硬件之间的接口界面。用户主要是通过软件与计算机进行交流。软件是计算机系统设计的重要依据。为了方便用户,为了使计算机系统具有较高的总体效用,在设计计算机系统时,必须通盘考虑软件与硬件的结合,以及用户的要求和软件的要求。软件的正确含义应该是:(1)运行时,能够提供所要求功能和性能的指令或计算机程序集合(2)程序能够满意地处理信息的
- 计算机导论(5)
weixin_45886926
1.软件1.软件的定义软件是用户与硬件之间的接口用户主要是通过软件与计算机进行交流的。从传统意义上讲,软件是计算机系统中与硬件相互依存的一个部分(非硬件部分)。简单认为:软件=程序+数据+文档(程序是按照事先设计的功能和性能要求执行的指令序列;数据包括使程序能正常运行的参数及程序运行中所处理的对象;文档是与程序开发、维护和使用有关资料的总称)2.软件的分类1.按软件的功能分类(1)系统软件:系统软
- 计算机导论课后总结四
别急啊
软件的定义软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。它是包括程序、数据及相关文档的集合。软件与硬件的层次关系图软件的分类按软件功能分类系统软件和应用软件。1.系统软件有操作系统、支撑软件操作系统比如Windows、MacOS、Linux等。支撑软件比如Microsoft公司的Studio、NET等。按软件的规模分类有微型、小型、中型、大型、基大型和极大型。按软件的工作方式分
- 计算机导论课后总结-4
cacia C
计算机导论
第三章走进软件3.1软件的定义■软件是用户与硬件之间的接口,用户主要是通过软件与计算机进行交流的。■从传统意义上讲,软件是计算机系统中的非硬件成分。■软件与硬件是相互配合的。3.2软件的分类3.2.1按软件的功能分类可分为系统软件和应用软件。■其中系统软件分为操作系统和支撑软件。操作系统有比如Windows、MacOS、Linux等。支撑软件有比如Microsoft公司的Studio、NET等。■
- [源码和文档分享]基于c++图形化界面的clock页面置换算法
ggdd5151
一、课程设计目的《操作系统》是一门重要的专业基础课,是涉及较多硬件知识的计算机系统软件课程。在计算机软硬件课程的设置上,它起着承上启下的作用。操作系统对计算机系统资源实施管理,是所有其他软件与计算机硬件的唯一接口,所有用户在使用计算机时都要得到操作系统提供的服务。操作系统课程设计的主要任务是研究计算机操作系统的基本原理和算法,掌握操作系统的进程管理、存储管理、文件管理和设备管理的基本原理与主要算法
- rust的指针作为函数返回值是直接传递,还是先销毁后创建?
wudixiaotie
返回值
这是我自己想到的问题,结果去知呼提问,还没等别人回答, 我自己就想到方法实验了。。
fn main() {
let mut a = 34;
println!("a's addr:{:p}", &a);
let p = &mut a;
println!("p's addr:{:p}", &a
- java编程思想 -- 数据的初始化
百合不是茶
java数据的初始化
1.使用构造器确保数据初始化
/*
*在ReckInitDemo类中创建Reck的对象
*/
public class ReckInitDemo {
public static void main(String[] args) {
//创建Reck对象
new Reck();
}
}
- [航天与宇宙]为什么发射和回收航天器有档期
comsci
地球的大气层中有一个时空屏蔽层,这个层次会不定时的出现,如果该时空屏蔽层出现,那么将导致外层空间进入的任何物体被摧毁,而从地面发射到太空的飞船也将被摧毁...
所以,航天发射和飞船回收都需要等待这个时空屏蔽层消失之后,再进行
&
- linux下批量替换文件内容
商人shang
linux替换
1、网络上现成的资料
格式: sed -i "s/查找字段/替换字段/g" `grep 查找字段 -rl 路径`
linux sed 批量替换多个文件中的字符串
sed -i "s/oldstring/newstring/g" `grep oldstring -rl yourdir`
例如:替换/home下所有文件中的www.admi
- 网页在线天气预报
oloz
天气预报
网页在线调用天气预报
<%@ page language="java" contentType="text/html; charset=utf-8"
pageEncoding="utf-8"%>
<!DOCTYPE html PUBLIC "-//W3C//DTD HTML 4.01 Transit
- SpringMVC和Struts2比较
杨白白
springMVC
1. 入口
spring mvc的入口是servlet,而struts2是filter(这里要指出,filter和servlet是不同的。以前认为filter是servlet的一种特殊),这样就导致了二者的机制不同,这里就牵涉到servlet和filter的区别了。
参见:http://blog.csdn.net/zs15932616453/article/details/8832343
2
- refuse copy, lazy girl!
小桔子
copy
妹妹坐船头啊啊啊啊!都打算一点点琢磨呢。文字编辑也写了基本功能了。。今天查资料,结果查到了人家写得完完整整的。我清楚的认识到:
1.那是我自己觉得写不出的高度
2.如果直接拿来用,很快就能解决问题
3.然后就是抄咩~~
4.肿么可以这样子,都不想写了今儿个,留着作参考吧!拒绝大抄特抄,慢慢一点点写!
- apache与php整合
aichenglong
php apache web
一 apache web服务器
1 apeche web服务器的安装
1)下载Apache web服务器
2)配置域名(如果需要使用要在DNS上注册)
3)测试安装访问http://localhost/验证是否安装成功
2 apache管理
1)service.msc进行图形化管理
2)命令管理,配
- Maven常用内置变量
AILIKES
maven
Built-in properties
${basedir} represents the directory containing pom.xml
${version} equivalent to ${project.version} (deprecated: ${pom.version})
Pom/Project properties
Al
- java的类和对象
百合不是茶
JAVA面向对象 类 对象
java中的类:
java是面向对象的语言,解决问题的核心就是将问题看成是一个类,使用类来解决
java使用 class 类名 来创建类 ,在Java中类名要求和构造方法,Java的文件名是一样的
创建一个A类:
class A{
}
java中的类:将某两个事物有联系的属性包装在一个类中,再通
- JS控制页面输入框为只读
bijian1013
JavaScript
在WEB应用开发当中,增、删除、改、查功能必不可少,为了减少以后维护的工作量,我们一般都只做一份页面,通过传入的参数控制其是新增、修改或者查看。而修改时需将待修改的信息从后台取到并显示出来,实际上就是查看的过程,唯一的区别是修改时,页面上所有的信息能修改,而查看页面上的信息不能修改。因此完全可以将其合并,但通过前端JS将查看页面的所有信息控制为只读,在信息量非常大时,就比较麻烦。
- AngularJS与服务器交互
bijian1013
JavaScriptAngularJS$http
对于AJAX应用(使用XMLHttpRequests)来说,向服务器发起请求的传统方式是:获取一个XMLHttpRequest对象的引用、发起请求、读取响应、检查状态码,最后处理服务端的响应。整个过程示例如下:
var xmlhttp = new XMLHttpRequest();
xmlhttp.onreadystatechange
- [Maven学习笔记八]Maven常用插件应用
bit1129
maven
常用插件及其用法位于:http://maven.apache.org/plugins/
1. Jetty server plugin
2. Dependency copy plugin
3. Surefire Test plugin
4. Uber jar plugin
1. Jetty Pl
- 【Hive六】Hive用户自定义函数(UDF)
bit1129
自定义函数
1. 什么是Hive UDF
Hive是基于Hadoop中的MapReduce,提供HQL查询的数据仓库。Hive是一个很开放的系统,很多内容都支持用户定制,包括:
文件格式:Text File,Sequence File
内存中的数据格式: Java Integer/String, Hadoop IntWritable/Text
用户提供的 map/reduce 脚本:不管什么
- 杀掉nginx进程后丢失nginx.pid,如何重新启动nginx
ronin47
nginx 重启 pid丢失
nginx进程被意外关闭,使用nginx -s reload重启时报如下错误:nginx: [error] open() “/var/run/nginx.pid” failed (2: No such file or directory)这是因为nginx进程被杀死后pid丢失了,下一次再开启nginx -s reload时无法启动解决办法:nginx -s reload 只是用来告诉运行中的ng
- UI设计中我们为什么需要设计动效
brotherlamp
UIui教程ui视频ui资料ui自学
随着国际大品牌苹果和谷歌的引领,最近越来越多的国内公司开始关注动效设计了,越来越多的团队已经意识到动效在产品用户体验中的重要性了,更多的UI设计师们也开始投身动效设计领域。
但是说到底,我们到底为什么需要动效设计?或者说我们到底需要什么样的动效?做动效设计也有段时间了,于是尝试用一些案例,从产品本身出发来说说我所思考的动效设计。
一、加强体验舒适度
嗯,就是让用户更加爽更加爽的用你的产品。
- Spring中JdbcDaoSupport的DataSource注入问题
bylijinnan
javaspring
参考以下两篇文章:
http://www.mkyong.com/spring/spring-jdbctemplate-jdbcdaosupport-examples/
http://stackoverflow.com/questions/4762229/spring-ldap-invoking-setter-methods-in-beans-configuration
Sprin
- 数据库连接池的工作原理
chicony
数据库连接池
随着信息技术的高速发展与广泛应用,数据库技术在信息技术领域中的位置越来越重要,尤其是网络应用和电子商务的迅速发展,都需要数据库技术支持动 态Web站点的运行,而传统的开发模式是:首先在主程序(如Servlet、Beans)中建立数据库连接;然后进行SQL操作,对数据库中的对象进行查 询、修改和删除等操作;最后断开数据库连接。使用这种开发模式,对
- java 关键字
CrazyMizzz
java
关键字是事先定义的,有特别意义的标识符,有时又叫保留字。对于保留字,用户只能按照系统规定的方式使用,不能自行定义。
Java中的关键字按功能主要可以分为以下几类:
(1)访问修饰符
public,private,protected
p
- Hive中的排序语法
daizj
排序hiveorder byDISTRIBUTE BYsort by
Hive中的排序语法 2014.06.22 ORDER BY
hive中的ORDER BY语句和关系数据库中的sql语法相似。他会对查询结果做全局排序,这意味着所有的数据会传送到一个Reduce任务上,这样会导致在大数量的情况下,花费大量时间。
与数据库中 ORDER BY 的区别在于在hive.mapred.mode = strict模式下,必须指定 limit 否则执行会报错。
- 单态设计模式
dcj3sjt126com
设计模式
单例模式(Singleton)用于为一个类生成一个唯一的对象。最常用的地方是数据库连接。 使用单例模式生成一个对象后,该对象可以被其它众多对象所使用。
<?phpclass Example{ // 保存类实例在此属性中 private static&
- svn locked
dcj3sjt126com
Lock
post-commit hook failed (exit code 1) with output:
svn: E155004: Working copy 'D:\xx\xxx' locked
svn: E200031: sqlite: attempt to write a readonly database
svn: E200031: sqlite: attempt to write a
- ARM寄存器学习
e200702084
数据结构C++cC#F#
无论是学习哪一种处理器,首先需要明确的就是这种处理器的寄存器以及工作模式。
ARM有37个寄存器,其中31个通用寄存器,6个状态寄存器。
1、不分组寄存器(R0-R7)
不分组也就是说说,在所有的处理器模式下指的都时同一物理寄存器。在异常中断造成处理器模式切换时,由于不同的处理器模式使用一个名字相同的物理寄存器,就是
- 常用编码资料
gengzg
编码
List<UserInfo> list=GetUserS.GetUserList(11);
String json=JSON.toJSONString(list);
HashMap<Object,Object> hs=new HashMap<Object, Object>();
for(int i=0;i<10;i++)
{
- 进程 vs. 线程
hongtoushizi
线程linux进程
我们介绍了多进程和多线程,这是实现多任务最常用的两种方式。现在,我们来讨论一下这两种方式的优缺点。
首先,要实现多任务,通常我们会设计Master-Worker模式,Master负责分配任务,Worker负责执行任务,因此,多任务环境下,通常是一个Master,多个Worker。
如果用多进程实现Master-Worker,主进程就是Master,其他进程就是Worker。
如果用多线程实现
- Linux定时Job:crontab -e 与 /etc/crontab 的区别
Josh_Persistence
linuxcrontab
一、linux中的crotab中的指定的时间只有5个部分:* * * * *
分别表示:分钟,小时,日,月,星期,具体说来:
第一段 代表分钟 0—59
第二段 代表小时 0—23
第三段 代表日期 1—31
第四段 代表月份 1—12
第五段 代表星期几,0代表星期日 0—6
如:
*/1 * * * * 每分钟执行一次。
*
- KMP算法详解
hm4123660
数据结构C++算法字符串KMP
字符串模式匹配我们相信大家都有遇过,然而我们也习惯用简单匹配法(即Brute-Force算法),其基本思路就是一个个逐一对比下去,这也是我们大家熟知的方法,然而这种算法的效率并不高,但利于理解。
假设主串s="ababcabcacbab",模式串为t="
- 枚举类型的单例模式
zhb8015
单例模式
E.编写一个包含单个元素的枚举类型[极推荐]。代码如下:
public enum MaYun {himself; //定义一个枚举的元素,就代表MaYun的一个实例private String anotherField;MaYun() {//MaYun诞生要做的事情//这个方法也可以去掉。将构造时候需要做的事情放在instance赋值的时候:/** himself = MaYun() {*
- Kafka+Storm+HDFS
ssydxa219
storm
cd /myhome/usr/stormbin/storm nimbus &bin/storm supervisor &bin/storm ui &Kafka+Storm+HDFS整合实践kafka_2.9.2-0.8.1.1.tgzapache-storm-0.9.2-incubating.tar.gzKafka安装配置我们使用3台机器搭建Kafk
- Java获取本地服务器的IP
中华好儿孙
javaWeb获取服务器ip地址
System.out.println("getRequestURL:"+request.getRequestURL());
System.out.println("getLocalAddr:"+request.getLocalAddr());
System.out.println("getLocalPort:&quo
 是电子的质量,
是电子的质量,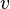 是其速度。假如光的频率太小的话,那么它无法使得电子获得足够的逸出功。这时,不论光强有多大,照射时间有多长,都不会发生光电效应。而当入射光的频率高于极限频率时,即使光不够强,当它射到金属表面时也会观察到光电子发射。
是其速度。假如光的频率太小的话,那么它无法使得电子获得足够的逸出功。这时,不论光强有多大,照射时间有多长,都不会发生光电效应。而当入射光的频率高于极限频率时,即使光不够强,当它射到金属表面时也会观察到光电子发射。