Nature子刊:微生物系统的功能与功能冗余

【论文题目】Function and functional redundancy in microbial systems
【期刊名称】Nature Ecology & Evolution,2018,2:936-943
【全文链接】https://doi.org/10.1038/s41559-018-0519-1
【第一作者】Stilianos Louca
【通讯作者】Stilianos Louca
【作者单位】哥伦比亚大学
摘要
微生物群落常常表现出惊人的分类(物种)多样性,这就引发了关于物种共存的机制、以及这种多样性在群落功能中的作用问题讨论。一方面,许多分类地位不同、共存微生物可以执行相同的代谢功能,这种功能冗余与期望物种本该占据的不同代谢生态位形成鲜明对比。另一方面,执行每个功能的分类单元的特征,在空间或时间上会发生很大变化,但对功能的影响却很小,这种分类组成变化常常被认为是等价生物体之间生态漂变的结果。在这里,我们整合了从两种模式中涌现出来的新范例,将微生物系统的功能、功能冗余和分类联系起来。我们的结论是,这两种模式不太可能是生态漂变的结果,而是开放式微生物系统不可避免的涌现属性,主要是由生物相互作用、及其环境与空间过程造成的。
前言
微生物是地球上最古老、最多样以及存在最广泛的生命形式。1克土壤中含有数千种微生物。微生物的代谢和生物合成多功能性同样令人印象深刻:已经发现的原核蛋白质编码的基因数量比所有动植物的总和都要多好几个数量级。微生物代谢途径驱动着多数生态系统中的大部分元素循环,在数十亿年的时间里塑造了地球表面的化学物质。然而,我们对微生物系统(微生物群落和耦合的非生物物理化学过程)的机理理解仍处于初级阶段。巨大的微生物多样性使建立微生物系统模型、解释群落在空间和时间上的变异模式面临着巨大挑战。此外,生态系统生态和生物地球化学中的许多问题需要了解微生物代谢功能的变化,而不仅仅是物种分类组成。
不管微生物的多样性有多高,大多数主要的生物地球化学反应还是由一系列有限的能量转换代谢途径所驱动,每种途径都存在于多种微生物进化分支中。群落功能谱研究(根据代谢功能描述群落)可以将微生物系统简化到数学模型允许的水平,从而揭示整个环境梯度中的群落结构模式。最近进行了一系列研究,从土壤到海洋和人类肠道等多种环境中,研究结果表明,代谢功能与环境因素之间存在强烈的耦合关系。许多情况下,在特定地点和时间,环境因素与物种分类组成发生解耦合。对各种代谢功能所涉及的微生物多样性的量化分析还表明,在多种功能方面,群落表现出高度的“功能冗余”,即每个代谢功能可由多种能够共存、分类学上不同的生物体来执行。目前对这些模式的含义和理解存在许多困惑,然而,对它们的合理解释为阐述控制微生物群落组成和功能的机制至关重要。在本文中,我们对这些模式进行了说明,并讨论了其中有说服力的范例,在塑造微生物系统时将功能、功能冗余和分类单元的作用结合起来。
解析微生物系统的功能和分类
第一次关于微生物群落的比较宏基因组调查表明,功能组成(就群落中发现的基因而言)与取样环境的类型(海水、土壤等)高度相关,表明特定的功能对环境有所选择。随后对不同人类宿主肠道微生物群落的比较表明,不同宿主之间微生物群落的分类组成差异很大,而它们的群落基因含量却高度保守(变化不大)。同样,在一项对大型藻类澳洲石莼(Ulva australis)的细菌群落调查中,群落构建似乎是基于功能基因而不是物种进行的。这些发现表明,不同的微生物组合表现出由环境选择的相似的群落基因组成谱。与本文提出的观点一致的是,对凤梨科植物叶“槽”内细菌和古细菌群落的最新研究发现,凤梨科植物群落的功能组成(就涉及各种能量转换功能的基因来说;图1c、d)高度保守。相反,不论使用何种分辨率进行分类(上升达到纲分类级别;图1a、b),与每个功能组相关的分类单元(即能够执行特定的代谢功能)在凤梨科植物群落之间的变化很大。因此,功能群内的分类组成必须是由不同于群落功能结构形成的附加因素所产生,即分类组成和功能组成(遗传潜力)呈现“解耦合”。在生物反应器(如脱氮或产甲烷)的实验中,反复观察到各种代谢功能与群落分类单元组成之间相似的解耦合现象,在生物反应器中,随着时间的推移,群落分类组成的高度变化伴随着稳定的生物反应器性能。下面,我们将讨论促进这种常见现象的条件和机制。
![]()
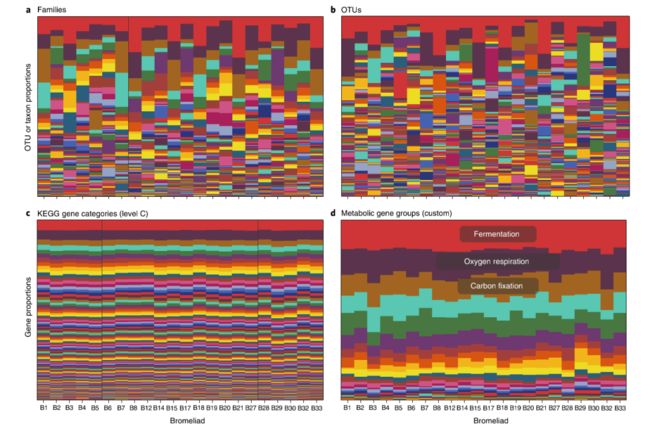
图1. 微生物群落中基因组成与分类组成的解耦合
a、细菌和古细菌科水平的分类单元相对丰度;b、细菌和古细菌株系水平可操作分类学单位(16S rRNA基因99%相似性)的相对丰度,样品来自巴西朱鲁巴蒂巴国家公园的22个类似的凤梨科植物叶,每一列代表一种凤梨科植物,每一种颜色代表一个分类单元。
c、基于京都基因与基因组百科全书(KEGG)划分的基因组成;d、自定义的代谢基因组成。每一列代表一个样本,每一种颜色代表一个基因组。值得注意的是,与相对保守的宏基因组组成相比,凤梨科植物的微生物分类组成更为多变。
稳定的功能组成和多样分类组成之间的鲜明对比,在之前的研究中已经有所发现,反映了许多功能和系统发育之间的弱相关关系。事实上,很大一部分代谢功能并不是单系的,也就是说,没有任何一个分支是这些功能的唯一代表。因此,虽然有机体在原则上系统进化地位决定了它的代谢能力(具有足够的分辨率和/或性状保守性),但反之却不一定成立,即代谢能力不一定能代表特定的分支(值得注意的是,有氧光合作用是一个例外)。功能或基因组流线型的适应性丧失、趋同演化和水平基因转移都改变了许多性状的系统发育信号。水平基因转移还导致基因组内性状的低遗传连锁现象,从而导致基因组间性状的重新分配。比如,一些大肠杆菌菌株在其蛋白质基因编码中重叠率不到40%。功能保守的系统发育尺度在不同功能之间存在很大差异,对于单一功能的系统发育保守性也可能在不同的进化枝之间发生变化(图2a,b)。例如,由脱硫杆菌、脱硫盐生菌和脱硫菌等可培养的微生物都具有利用硫酸盐的能力,而利用硫酸盐能力只有古球菌属中的一个进化枝才有。由于既定的代谢功能,存在不同层次的不同分支中并被保存,因此缺少一个分类学上固定的分辨率,在这个分类学分辨率上,分类单元要么总是表现出这种功能,要么永远不表现出这种功能。因此,没有单一的分类学解决方案,其中分类学的变化明确反映了功能的变化,某些功能的环境选择(如好氧菌对氧气的选择)明确转化为对特定分类单元的选择。

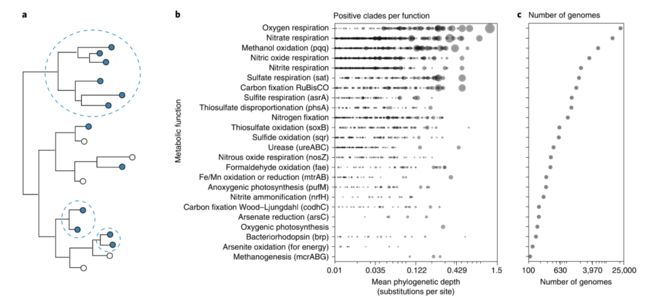
图2. 系统发育保守性在功能和进化枝之间存在差异
a、系统发育树的示意图,其中填充和开放的圆圈分别表示特定功能的存在和缺失。根据树中的位置,功能可以存在深或浅的分支(虚线圆圈)中。
b、在各种代谢功能中呈阳性的原核生物分支(即提示中功能≥95%),用圆圈表示(一个圆圈代表每个功能的阳性进化枝)。根据进化枝的平均系统发育深度(对16S rRNA基因中每个位点替换进行测量)将圆圈定位在水平轴上。较大的圆对应于包含更多备注提示的进化枝(对数刻度)。大多数功能在许多不同深度和大小的进化枝中都是保守的,而有氧光合作用是一个明显的例外。因此,对于大多数功能,不存在分类上的分辨率,分类单元要么总是表现出这种功能,要么从不表现出这种功能。
c、非冗余原核生物基因组的数量(即,具有唯一的NCBI分类单元ID),从NCBI RefSeq下载,发现展示每个功能。
图b和c基于在大约59000个全基因组测序微生物中检测到的基因(括号中列出了单个基因)。
对于某些功能与特定分类组成完全解耦合似乎是不可避免的,因为同样的功能可以由其他分类单元来执行(图2c)。营养物质的供应速率、光照度、地球化学梯度、环境迁移过程、以及生物体之间的化学计量平衡可以强烈地限制反应速率,而代谢通道的能量产出进一步影响功能群的增长速率。虽然每个功能只能由某些分类单元执行,但上述因素在特定情况下可能对这些分类单元中的某些分类单元所执行的功能施加较小的控制。与此相应的是,在时间或空间上,大部分生物化学通量速率,对功能群内的分类变化表现出低敏感性。为支持这一解释,对土壤进行的全球生物地理学研究发现,非生物土壤特性在很大程度上解释了氮循环途径中的丰度变化,但对相应功能群内的物种分类组成解释不够充分。对全球海洋普遍的代谢功能也进行了类似的研究。同样地,最近的meta分析发现,除了环境变量以外,将群落分类单元组成作为碳、氮循环过程速率预测因子的研究中,只有29%的研究提高了预测能力,将R2的平均值从0.56增加到0.65。哪些功能受到环境的强烈控制,从而对生物分类变化不太敏感,这取决于生态系统的类型,特别是通过氧化还原失衡获取能量以及物理化学边界条件。在实验中,广泛分布的功能,如呼吸作用、总碳分解代谢和生物量产生,比特定化合物的降解等狭义功能更能抵抗群落物种分类单元组成或多样性的变化。从功能上解释该模式的原因是,广泛的功能可能相对更为冗余,从而能更好地抵御由生物或非生物干扰引起的分类组成变化。热力学上有利于分解代谢途径的线性末端对分类变化的敏感性也可能低于个别中间步骤,这些中间步骤能够通过其他途径进行代谢。例如,持续补充葡萄糖的产甲烷生物反应器模型表明,通过“替代”分解代谢途径的相对通量速率(例如从葡萄糖到挥发性脂肪酸,最终到甲烷的各种替代途径)在分类组成变化上可能不如甲烷整体的产生率稳定(图3)。
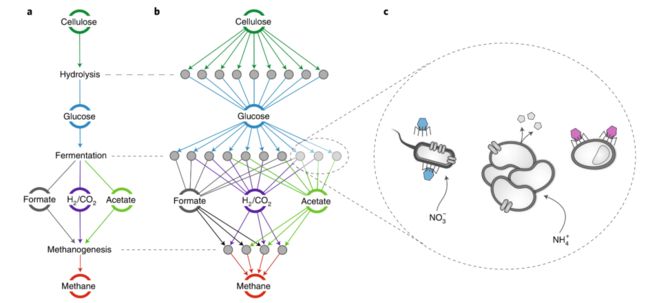
图3. 产甲烷群落的功能冗余示意图
a、微生物群落在产甲烷纤维素生物反应器中,使纤维素分解为甲烷的典型代谢网络。圆圈代表底物或产物,边缘颜色表示相关底物。
b、展开每个分解代谢步骤,显示出多个不同的有机体进行同一反应。灰色小圆圈代表不同的群体基因组。
聚焦将葡萄糖分解为醋酸盐的三个冗余有机体。通过所利用的底物类型以外的性状差异来实现实际生态位分化与共存,可能包括对不同噬菌体(蓝色和紫色)的敏感性、不同的捕食策略、颗粒附着和生物被膜形成,使用不同的氮库(硝酸盐与氨),以及不同抗生素(小五边形)的产生及其耐药性。
已经有一些研究观察到群落功能和群落分类组成之间的强相关性,例如沿着强氧化还原梯度。我们强调,当环境条件变化时,除了功能群内发生的分类组成变化外,对特定代谢功能的选择通常会导致群落分类单元组成的变化。因此,当在空间或时间上对群落进行比较时,群落功能和群落分类单元组成之间的相关性将取决于针对特定功能选择的机制与导致功能组内部变化的机制之间的相对重要性(接下来将进行讨论),以及这些功能的系统发育分布。
我们指出,功能共同体结构原则上可以定义为任意一组功能(功能的选择决定了观察到的时空格局),尽管有对能量转换代谢功能进行特别关注,但这些功能涉及主要的元素循环或具有特定工业重要意义。我们还认为,一些作者定义了“功能反应群”(即对特定环境因子有相似反应的生物体群),并将其与“功能效应群”区分开来(即对特定生态系统功能有相似作用的生物体群)。然而,在这里我们避免使用这个术语,部分原因是(代谢)功能组,从这个角度来看,既可以是效应群、也能是反应群。此外,如前文所述,代谢功能组内的代谢功能和分类变化构成了许多微生物系统的互补面和分离面,能深入了解显著不同的过程。
功能冗余在微生物系统中普遍存在
很大一部分代谢基因在地球早期阶段已经出现,正如上文所讨论的,随着地质时间的推移,这些基因传播到多个微生物进化枝中去。现今,在全球范围内,大多数代谢功能由大范围的现存微生物类群执行。更为突出的是,即使是在局域尺度上,与每一种代谢功能相关的分类单元计数,无论是通过对宏基因组序列进行分类,还是通过分类单元的功能进行分类,往往都会发现能够执行类似代谢功能的多种不同生物共存。例如,数百种能够进行氢氧化的微生物在地下水中共存、数百种产氧自养生物在海洋表面共存。在海底含水层中,有几十个基因组能够通过氧化硫化物来获取能量,至少有15个基因组能够进行反硝化作用。在产甲烷反应器中,水解纤维素可由数十种不同的微生物同时进行。在硝化生物反应器中,通常有多个氨氧化细菌共存,并且随着时间的推移,呈现出不同的相对丰度变化。功能冗余是许多微生物系统的共同特征。也就是说,很明显,任何一个特定系统的功能冗余程度都取决于所考虑的功能。例如,在阳光充足和富含氧的海洋表面,光能自养和有氧呼吸作用通常比硫酸盐呼吸作用和甲烷生成更为冗余。
群落功能结构(以及功能冗余)原则上可以在不同的细节层次上进行定义,例如基于反应动力学进一步区分功能定义。一些研究者认为,只有当生物体在生态位高度相似性,从而能够很容易地相互替换时,这些生物体在功能上才是冗余的,尽管他们承认这一标准在实践中很难满足。其他研究者认为,在特定的相同环境条件下,如果生物体能够以同一速率执行某一项功能,那么它们就是冗余的。后一个要求在实践中很难检验,并且测序数据很少考虑超出催化反应能力类型推测的酶动力学。这种定义的实用性还有一个实际限制,即一个种群的代谢活动依赖于群体的整体状态,例如是否存在交叉互饲、噬菌体或杀菌素。此外,大部分过程的速率在很大程度上受到环境物理、化学性质的限制,如沉积物中的空间迁移速率或生物反应器中的底物供应速率。因此,具有不同反应动力学的分类单元中的种群,诱导形成不同或相同的生化通量速率,这取决于详细的环境条件(参数设置)以及当前的群落状态。因此,我们认为,与其他研究中更严格的定义相比,功能冗余的定义更具有实际意义,该定义仅仅表示多个不同生物体执行某一特定功能的能力(BOX1术语表)。例如,功能冗余(即本文中提到的)通常与功能对环境干扰的稳定性联系在一起,有助于深入了解重要的群落过程,如下文中所述。
BOX 1 术语表
功能群
分类单元能够执行特定的生化功能,比如根据其基因含量进行分类。
(群落)功能丰富度
重要的生化功能或基因的存在数量。
功能冗余(相对于特定功能而言)
能够执行相同的重要生化功能的、多个共存的分类单元或基因组。
(群落)功能结构
各种重要功能群或与重要功能基因的相对丰度。
生态漂变
有限种群中,出生-死亡事件的随机性导致种群相对大小的波动。
(生态系统)代谢生态位
生物体利用特定的代谢途径(例如,H2/CO2生成甲烷)或半反应(例如,使用特定的电子受体进行呼吸作用)获得生长所需能量的能力。
(群落构建)代谢生态位效应
生物体选择能够利用特定代谢生态位的机制。这些机制可能包括光合作用中光的可用性,或呼吸作用中作为电子受体的硫酸盐的可用性。
微生物系统
一个微生物群落,在细胞外环境中的代谢产物以及双向耦合的、非生物、物理化学过程,包括物理转运过程和非生物化学反应。与“生态系统”类似,但关注的是微生物成员,而不是宏观食物网。
促进功能冗余的机制
长期以来,人们在微生物群落中观察到能量转换代谢途径存在高度的功能冗余。例如,几乎所有植物都有一个共同的代谢生态位,因为它们全部是能够产氧的自养生物。在微生物和类似宏观生物中,功能冗余表明,除了单纯提供不同能源之外,附加因素也必须要控制多样性。的确,Tilman经典竞争理论声称,在状态稳定、充分混合的系统中,任何一种特定资源(如电子供体或受体)最多只能限制一个持久种群。该种群将在最低资源水平下保持一定规模大小的稳定,因为其他所有种群要么在竞争中胜出,要么受到另一资源的限制。尽管认为状态稳定和充分混合是一种理想情况,但Tilman竞争理论提供了一个基准(最低期望),能够与观察到的多样性进行比较。对于大型生物群落来说,一个物种在有限的资源中持续存在的理论预期与观察到的生命多样性之间有明显的脱节(不一致),这一现象已经通过多种方式进行了解释。首先,在资源受限的特性或环境条件下的时空异质性,再加上物种之间的响应差异,可能会形成多重生态位。第二,竞争排斥可能被诸如捕食之类的生物相互作用所破坏,或者由于区域库中的扩散而被抵消。重要的是,物种可能在参与资源竞争的性状和参与环境耐受性、抗捕食性或扩散能力性状之间进行权衡。
与大型生物体相似,微生物群落的功能冗余可能是由其他生态位轴的分化促进的,而不仅仅是代谢资源。这些代谢资源包括生物体对环境干扰的响应差异、对颗粒的依附策略差异、营养梯度和食物颗粒的趋药性策略差异、特定多糖(例如,藻酸盐)裂解酶基因的数量和类型差异、营养浓度的波动与不同的生长动力学结合、不同微量营养素的限制以及噬菌体和原生动物的捕食作用。例如,尽管我们不清楚噬菌体对微生物群落的确切影响,但是在获取养分和抵抗噬菌体侵袭之间进行权衡,能够使竞争菌实现共存。非传递性竞争动态,即多对共同的竞争物种之间没有明确的赢家,也能在抗生素抗性中发挥作用。新陈代谢重叠的微生物以我们目前能够识别的多种方式进行生态分化,因此群落组合发生在高维度(多因子)空间内。事实上,最近对微生物基因组进行的基因注释研究工作,发现了成千上万的基因簇,其中大部分是尚未表征明确的功能。根据这些研究,在开放的微生物系统中,功能冗余似乎是不可避免的结果,在这些系统中,多样性不受低迁移率的限制。
仅仅根据代谢能力来评估生物体的代谢生态位时,必须多加注意,例如从生物体的基因组推断其代谢生态位。具有相似代谢系统(“基础”代谢生态位)的种群专食不同的营养物质,因此在转录水平可能显示出分离的“实际”生态位。特别是,即便只有少数几个成员活跃地执行代谢功能,功能群也会表现出高度冗余,这是因为一部分成员会以替代模式获取能量,而其他成员则并不活跃。某一特定种群的代谢功能通常取决于环境条件以及其他群落成员的存在和活动。我们强调,即使一个群落中的生物体进行多功能代谢,前文讨论的经典竞争理论预测仍然适用。也就是说,在稳定状态下,共存生物体的数量不能超过限制至少一种生物生长的资源数量(包括代谢副产物)。例如,两个氢自养产甲烷菌可以在同一环境中共存,在稳定状态下,它们并不会受到同一氢气库的限制。非充分混合环境中的小尺度空间分离可能是一种共存机制。例如,具有类似营养偏好的生物体可以在不同的生物被膜上定殖并获取营养物质,从而能够在更大的尺度上实现共存。然而在这些情况下,重要的是,要认识到在不同的生物被膜上的种群不会竞争相同的营养库,因此会有不同的实际生态位。
功能冗余并不意味着中性
之前的一项研究假设,代谢生态位内的功能冗余可以反映竞争者近中性的共存。然而,如前面讨论的,共存微生物专食同一营养资源,不仅在酶效率和生长动力方面存在显著的差异,而且在特定条件下,影响其生长速率的其他性状也有所不同。虽然人们普遍承认功能群成员之间的差异,但是关于微生物群落组成的某些模式是否可以用中性过程来解释仍存在争议。类比于宏观生态的中性理论,有人提出了一个用于研究局域微生物群落组成的中性模型,这个模型仅仅基于随机迁移和生态漂变(有限种群中由出生/死亡事件的随机性引起的波动),但却忽略了物种形成,物种形成是宏观生态中性理论的一个常见过程。他们得出的结论是,随机迁移和生态漂变是形成原核生物群落的重要因素,特别是在代谢功能群中。在这项研究之后,中性模型被用来解释一部分不同环境中的微生物生物地理格局,这些环境包括动物肠道、土壤、生物反应器、树洞和生物被膜。还有人提出,功能群内的生态漂变可以解释一部分物种的时间更替,例如在生物反应器、地下水和在河流集水区中的物种随时间的更替。
我们强调,即使在封闭系统中,功能群内复杂或明显的分类组成的随机变化也不能与生态漂变混为一谈。事实上,生态漂变并不能有效地解释功能群内的分类组成更替,例如在生物反应器中观察到的随时间的更替。这是因为与选择过程相比,生态漂变在大种群规模和/或竞争者之间巨大的生态差异中,其重要性降低了。在生物反应器和大多数的自然环境中,细胞密度非常高(生物反应器中的细胞数量可达1013个/L),以至于选择过程明显优于生态漂变。事实上,中性的随机出生-死亡模型预测,即使在种群规模较小(104个细胞/L)的情况下,在由同等竞争者组成的群落中,相对稀有的生物体(1%的比例)仅通过生态漂变(世代时间为1天)达到30%的比例,平均需要1600多天。甚至假定其中一种生物的竞争优势较弱时(预期增长率提高5%),两个种群都严格遵循由竞争排斥预测的确定的轨迹(解释方差为0.98±0.02)。因此,在微弱的竞争差异下,生态漂变对种群轨迹的影响可以忽略不计。我们注意到上述模型参数是相当保守的。事实上,微生物种群通常包括104个以上的细胞,即使在恒定的环境条件下,观察到极其稀有的分类单元(少于0.1%的比例)在短短几周内取代之前占主导地位和代谢相似的分类单元也并不罕见。此外,即使是同一物种不同菌株也明显表现出不同的底物亲和力,或者对专门的噬菌体的敏感性不同。因此,竞争者在足够长的时间内具有相似的生长率,使生态漂变成为分类组成更替显著驱动因素的可能性极低。因此,尽管功能冗余(无论是在局域范围还是区域范围内)是功能群内分类组成更替的必要条件,但更替本身一般不能用生态漂变来解释。与这一预测相一致的是,最近对人体大规模的微生物群落分析发现,只有不到1%的群落符合Hubbel中性生物多样性理论。同样,一项对凤梨科植物微生物组的调查结果表明,尽管凤梨科微生物群落功能结构稳定、功能冗余度高、分类组成高度变化,但其功能群组合并非中性变化。即使在植物和动物生态学中,其种群规模远远小于典型的微生物群落规模,也缺少明确的证据能够表明生态漂变的强大作用(例如,与选择相比)。
由于生态漂变通常不能解释功能群内的分类组成更替,这种更替必然是由功能群成员之间的生态差异和潜在的扩散过程造成的。以往的研究确实表明,有限扩散是不同地点之间分类组成变化的一个重要来源,这是基于早期定殖者在群落演替过程中的随机系统发育结构,随着时间的推移,半开放培养阶段的分类丰富度逐渐增加,或者更常见的是,群落相似性随着地理距离的增加而衰退。然而,在此之后的研究仍没有形成定论,因为群落相似性的距离衰减也可能是由空间相关的环境异质性引起的。例如,对环境异质性的解释可用于阐明盐沼、全球海洋和凤梨之间的距离和微生物群落相异性之间的全部或大部分的相关关系。环境异质性是植物群落分类组成空间变化的主要原因之一,如果没有进行充分的环境因子测量,环境异质性一般很难排除。
在恒定条件下重复操作的生物反应器的实验中,微生物群落组成遵循复杂但却重复的轨迹,周期从几周到几个月不等。这表明,在没有明显环境变化的情况下,功能群内的分类组成更替可以由内在的和至少部分确定性的过程来驱动。这种内在过程可能包括“杀死获胜者”型噬菌体宿主相互作用,即专门的噬菌体反复诱导优势微生物种群的崩溃,目前这种机制的实验证据很少。提出的其他机制包括抗生素抗性、交叉互饲的快速演化和适应生态位构建。因此,每个物种都可能受到调节其瞬时生长率的生物和非生物因素组合的影响,即使其代谢能力与群落中的其他成员重叠。这些影响因素可能对频率有所依赖,还可能包括随机成分,例如基因突变或基因水平转移。在实践中,混沌种群动力学可能会模糊群落构建的确定性过程和随机性过程的区别。此外,在区域尺度范围内,不常见的扩散作用会增加群落组成的随机性,这不能仅仅通过内在动力学来解释。因此,即使清楚某一特定时刻所有的环境因素,也不可能完全预测群落分类单元组成。
结论
通常认为功能冗余是群落构建中性过程的一种表现,实际上它是具有特定代谢功能的微生物生态多样性的表现。功能冗余是开放微生物系统不可避免的涌现特征,当一个具有高维度特性空间被投射到一个低维的空间时,就会看到这种特征。因此,可以将功能冗余看作是多样性的部分测量,即功能群体内的多样性,这在数学上是对群体功能丰富度的补充,正如将功能群体内的分类组成视为对功能群体结构的补充。我们推测,开放微生物系统的功能冗余度可能是一个稳定的系统特征,这在很大程度上取决于环境类型和所考虑的功能。这一假设对于不断受到迁移影响的自然系统来说尤其正确,比如在开放的海洋中,物种迁移和当地物种灭绝之间的平衡可能决定生态时间尺度上的功能冗余。
根据功能的选择,区分群落功能结构和功能群内的组成可以对生物地球化学和群落构建机制有重要的了解。事实上,与能量转换相关的代谢途径可以与特定的环境因子和元素循环强烈耦合,同时与特定分类组成解耦合。在宏观生态学中也得到类似的观察结果,在描述群落结构的共位群、生命形式和生存策略方面已经有很长的研究历史,认为这些都类似于微生物中的代谢功能群。最近,甚至有人提出完全放弃物种方面的宏观群落模型,把重点放在功能性状上。将复杂的微生物群落简化为能量转化代谢功能性状的方式,同时研究与这些功能有关的功能冗余,这对微生物生态学也可能是一个富有成效的方法。
除了代谢生态位效应以外,即使某些代谢功能的活性是强烈保守的,一些其他的机制,如捕食和抗生素抗性,将会随着时间和空间变化调整微生物群落的分类组成。显然,群落水平功能与分类组成之间的这种明显的解耦合现象,并不是功能群内部随机生态漂变的简单结果。各种机制如何以及在何种条件下将产生这种解耦合现象,以及是什么决定了微生物系统功能冗余的程度,正在成为生态学研究的中迫切需要解答的关键问题。
翻译:马国琳
校对:任可欣、王文平
猜你喜欢
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature Cell专刊 肠道指挥大脑
系列教程:微生物组入门 Biostar 微生物组 宏基因组
专业技能:学术图表 高分文章 生信宝典 不可或缺的人
一文读懂:宏基因组 寄生虫益处 进化树
必备技能:提问 搜索 Endnote
文献阅读 热心肠 SemanticScholar Geenmedical
扩增子分析:图表解读 分析流程 统计绘图
16S功能预测 PICRUSt FAPROTAX Bugbase Tax4Fun
在线工具:16S预测培养基 生信绘图
科研经验:云笔记 云协作 公众号
编程模板: Shell R Perl
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流、快速解决科研困难,我们建立了“宏基因组”专业讨论群,目前己有国内外5000+ 一线科研人员加入。参与讨论,获得专业解答,欢迎分享此文至朋友圈,并扫码加主编好友带你入群,务必备注“姓名-单位-研究方向-职称/年级”。PI请明示身份,另有海内外微生物相关PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。

学习16S扩增子、宏基因组科研思路和分析实战,关注“宏基因组”
 点击阅读原文
点击阅读原文