4-8岁儿童EEG微状态研究:年龄和性别的影响
大规模神经网络的超快时空动态可以通过静息状态脑电图(EEG)微观状态来检验,这代表了随时间动态演化的同步神经活动的短暂时期。在成人中,四种典型的微观状态已被证明可以解释静息状态脑电图的大部分地形差异。它们的时间结构依赖于年龄、性别和状态,并且易受大脑病理状态的影响。然而,没有研究只评估了儿童早期-大脑快速发育的关键时期-脑电图微状态的空间和时间特性。本研究试图调查在103名4-8岁儿童的大样本中使用高密度脑电图记录的脑电图微观状态。利用数据驱动的k-means聚类分析,发现在成人人群中报告的四种典型微观状态已经存在于儿童早期。使用多元线性回归,我们证明了两种微观状态的时间动态与年龄和性别有关。源定位表明,注意力和认知控制相关的网络影响着年龄和性别依赖的微观状态的地形。这些新发现为用脑电图微观状态捕获的儿童的脑功能发育提供了独特的见解。
1.介绍
幼儿期是一个大脑快速发育和行为改变的时期。在儿童早期描述基于网络的大脑连接的发育特性可能有助于我们理解神经认知发育的规范和非典型模式。脑电图微观状态是头皮电位地形的模式,大约每60-120毫秒相互转换一次,反映了随着时间动态演化的同步神经活动的短暂时期。最近的研究表明,仅两分钟的脑电图数据就可以得到脑电图特性的可靠估计。然而,迄今为止很少有研究在儿童中使用这种方法,也没有项研究专门关注8岁以下的儿童。先前使用聚类方法对老年组进行的研究表明,四种微观状态——典型标记为A-D——解释了静息状态脑电图头皮电位地形图的大部分差异。同时进行的脑电图功能磁共振成像(fMRI)研究以及脑电图源成像表明,这些微观状态的空间模式类似于众所周知的静息状态网络(例如,微观状态A代表听觉,B代表视觉,C代表显著性,D代表注意网络)。现有的研究表明,微观状态之间的快速过渡反映了大规模功能网络的动态重组。重要的是,脑电图的毫秒时间分辨率允许量化关于每个微观状态的时间属性的独特信息,包括其全局解释方差(GEV;即,由给定微观状态解释的数据中总方差的百分比)、平均持续时间、它存在的时间百分比(即覆盖率)和每秒发生的频率。因此,先前的研究支持脑电图微状态分析作为一种测量基于网络的连接的时空特性的新方法。
年龄和性别对静息状态脑电图时空动态有影响,特别是微观状态C和D的持续时间和发生率。以往相关都没有利用源定位来确定每个微观状态下的大脑区域,且不分年龄确定了群体水平的微观状态,忽视了一些重要的儿童脑电图微观状态地形和时间动态的发育变化。本研究采用103例4-8岁儿童高密度脑电图,检测了脑电图微观状态及年龄和性别效应的时空特征。预估时间参数会随着年龄的增加而变化,并显示出性别差异。通过定位微观状态的来源发现,相比于运动和感觉网络的微观状态而言,更大的年龄和性别效应出现在参与高阶认知功能和网络整合的网络微观状态中。
2.方法
2.1 被试
被试是从杜克大学心理学和神经科学系和社区活动维护的数据库中招募的4-8岁的儿童。在筛选了323名儿童后,有249名符合条件。其中,171名儿童完成了他们的第一次研究访问。静息状态脑电图在数据收集时被添加到研究中。当脑电图是研究方案的一部分时,参与研究的合格儿童的数量为140人。32名儿童没有完成脑电图,因为不配合(例如,拒绝佩戴脑电图网)。总之,108名参与者在第一次或第二次研究访问中完成了脑电图测试,其中103名参与者提供了足够的分析数据。
2.2 脑电图数据采集和预处理
使用128通道 HydroCel Geodesic Sensor Net (Electrical Geodesics, Eugene, OR),1000赫兹采样,在线顶点参考。在整个范式中,阻抗保持在50千赫以下,其中包括8个1分钟的交替睁眼和闭眼的静息状态(即每种情况下的4分钟)。实验者给出的语言线索和通过E-Prime软件显示的文本指导参与者睁开眼睛或闭眼放松。本研究只分析了闭眼的情况。
离线预处理是使用MATLAB和EEGLAB执行的,使用https://github.com/DEEDLabEEG上提供的自定义脚本。位于脑电图网络外环上的通道被移除。数据被降采样到250 Hz,在40 Hz低通滤波,在1 Hz进行高通滤波。在60 Hz的剩余电线噪声使用CleanLine plugin进行衰减。坏信道-超过5秒的1)平坦,2)包含相对于它们信号的线噪声超过4个标准差,3)与基于附近信道的估计相关小于0.8-被去除。Artifact Subspace Reconstruction(ASR)删除了包含伪迹的部分数据。如果超过25%的信道功率超过7个标准差,则删除额外的数据周期。使用主成分分析(30个成分),并将ICA矩阵复制到全长数据中(即在ASR去除伪迹之前)。ICLabel插件删除眼动和肌动概率大于0.7组件。如果至少有10个通道的振幅大于1)100μV或小于−100 μV,或2)联合概率超过局部/全局阈值的三个标准差,则数据被分割为不重叠的一秒周期,并使用TBT插件删除。如果少于10个通道满足拒绝标准,则不删除该时代,但只为该时代插值该通道。最后,使用球形样条插值,所有通道重新引用到平均值。经过预处理和删除5名数据没有通过质量控制的被试后,103名被试的最小数据量为145秒(s)。为了减少不同参与者之间不同数据长度对进一步分析的潜在影响,所有参与者的数据被裁剪到前145秒。
2.3 微状态分析
使用Cartool进行微观状态分析,首先在个体水平,然后在组水平。在个体层面,对每个被试的脑电图数据应用空间滤波器,以去除地形异常值和平滑地形。对于每个参与者的脑电图数据,提取全局场功率(GFP)峰值的地形,代表最高信噪比的时间点。每个被试先前提取的GFP峰(覆盖每个参与者数据的99.9%)的有833个随机子样本的50个epoch被提交到极性不变修正的k-均值聚类分析,设置为重复50次,并为每个epoch识别1-12个地形簇。包含7个独立标准的元标准综合测量确定每个epoch的最优簇数,得到50个epoch,每个epoch由k个最优簇组成。在组水平上,在个体水平分析中确定的50个最优集群被合并为一组,得到5150组(103名被试×50组)。接下来,将每个由1500个随机抽样集(覆盖99.7%的集合)组成的100个epochs提交到极性不变修正的k-means聚类分析中,该分析设置为重复100次,并为每个epochs识别1-15个地形聚类。元准则确定每个epoch的最优簇数,从而得到100个epoch,每个epoch由k个最优簇组成。最后,将这100个集合组合并提交到最终的极性不变修正k-means聚类分析,该分析设置为重复100次,识别1-15个聚类地形图。元标准确定了簇的最佳数量,现在被称为组水平的微观状态。
这些微观状态被反馈到每个被试的原始空间过滤数据中,包括所有的数据点(而不仅仅是在GFP峰值处)。数据由GFP的中位数归一化,以解释由于颅骨电导率不同而导致的头皮电位的个体差异。反拟合包括计算组水平上的每个微观状态与每个参与者的每个个体数据点之间的空间相关性,从而将相关性最高的微观状态分配到该数据点。在计算相关性时,忽略了地图的极性。分配给一个微观状态的数据点的最小相关性为50%。回拟后,采用时间平滑(窗口半尺寸为32 ms,Besag因子10),去除不太可能的小段,将小于32 ms的段分成两半,前半段加到前段,后半段加到下段。回拟合过程产生了每个微观状态的GEV、持续时间、覆盖范围和发生率。
2.3.1 微观状态的源地定位
脑电图网络模板与MRI头部模型进行共配准。具有解剖约束的局部球形模型(LSMAC)通过估计每个电极下的头皮、颅骨、脑脊液和大脑的厚度,计算出了每个电极上的自适应局部球形模型。LORETA通过正则化对结果进行优化,考虑了背景脑电图噪声和结果的强制平滑性,并进行标准化,以纠正脑电图功率随时间的变化(这些程序在Cartool中自动实现)。偶极子的振幅被保存为标量,每个解点的正值,并在每个微观状态的时间点上取平均值。每个微状态的源图的阈值设置为被试激活的第95百分位以上。源图被转换为体积,并导入到功能神经图像分析(AFNI)程序中。在AFNI中,他们通过从每个源图中减去所有源图的平均值来突出显示微状态特定的源。
2.4 统计分析
在R中进行统计分析。计算4个单向、重复测量、等n/II型平方和、方差分析(ANOVA)模型,以比较微观状态之间每个时间参数(GEV、持续时间、覆盖率、发生)的平均值。使用箱线图确定极端异常值。如果违反了球形度的假设,则采用Greenhouse-Geisser correction。对于每一个具有统计学意义的方差分析,在微观状态之间进行了事后配对t检验,p值进行了24次比较的Benjamini-Hochberg-corrected。使用DABEST软件包计算了基于估计统计量的效应大小和置信区间。然后,以每个微观状态的时间参数为因变量,以平均为中心的年龄、虚拟编码性别(男性=−1,女性= 1)和年龄按性别交互项为自变量,分别进行多元线性回归。多变量异常值分别由每个回归的最小协方差决定去除。为了最小化I型错误率,对完整预测因子的整体显著性的综合F检验的p值进行了Benjamini-Hochberg-corrected-16次比较校正。通过检验预测微观状态时间参数的年龄斜率是否对每个性别都显著,来探索具有统计学意义的交互作用,并使用 Johnson-Neyman procedure来确定在哪个年龄(s)时,性别的简单斜率在p < .05时与零有显著差异。
3.结果
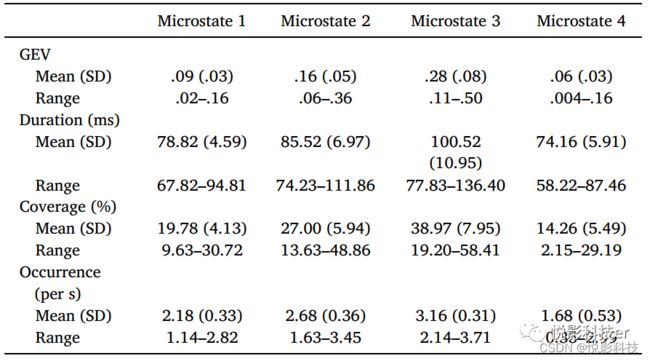
确定最佳微观状态数量的元标准显示了4个微观状态,分别标记为1-4(如图1)。在之前的文献中,微观状态1-3分别对应于典型的A-C。微观状态4的地形在以往的文献中不太一致;一些研究显示额中央最大值,而另一些研究显示后中央最大值。在微观状态回拟原数据中,平均3.40 s的数据(SD = 1.76 s)没有达到50%的最小空间相关阈值。对微观状态的时间参数的描述性统计数据见表1。
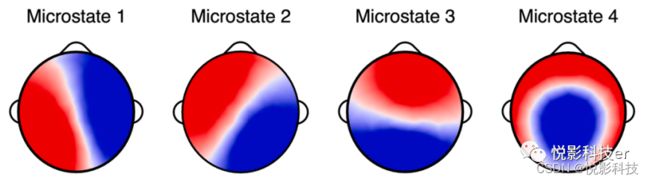
图1.四种微观状态。注:微观状态由极化不变聚类算法导出。
表1四种微观状态的时间参数的描述性统计图片
3.1 微观状态间时间参数均值差异
采用单因素重复测量方差分析持续时间、覆盖率和发生率的平均值,研究了四种微观状态之间GEV、持续时间、覆盖率和发生率的差异,排除异常值后模型结果如图2所示,同时还计算了基于估计统计的效应大小和95%ci。
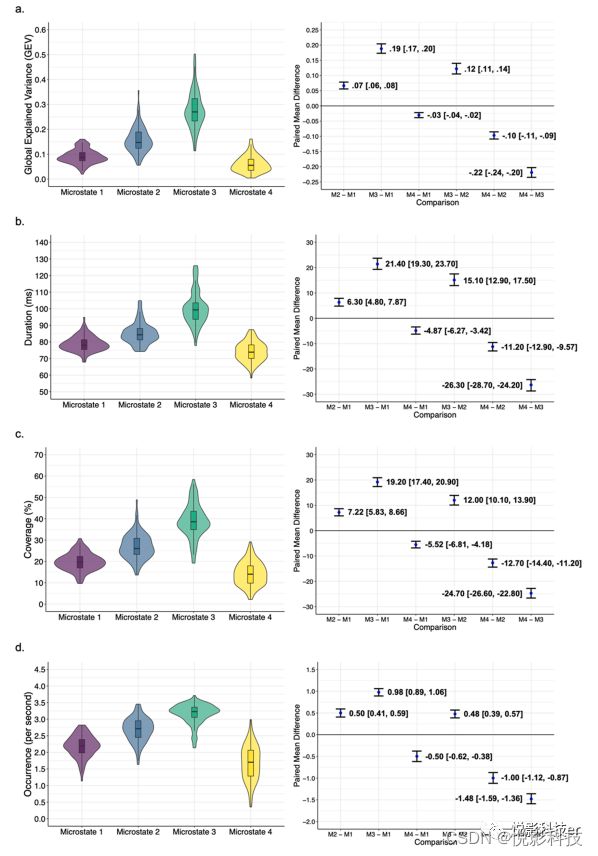
图2.小提琴图和箱形图(左)和成对的平均差异估计图(右)显示了不同微观状态之间的时间参数的平均值的差异。
不同微观状态之间的平均GEV、平均持续时间和平均覆盖率有显著性差异,事后检验都为,微态状态4<微观状态1 GEV<微观状态2<微观状态3。
3.2 年龄和性别对各微观状态的时间参数的影响
使用多元线性回归模型检验了每个微观状态的四个时间参数被性别与年龄交互作用、性别和年龄的预测程度,Benjamini-Hochberg-adjusted校正总体模型水平上进行了16次多重比较。为了便于可解释性,Johnson-Neyman intervals的年龄值以年为单位,并且仅在样本观察值范围内。排除异常值的模型的结果见表2。
表2 回归检验年龄和性别对每个微观状态的时间参数的影响。
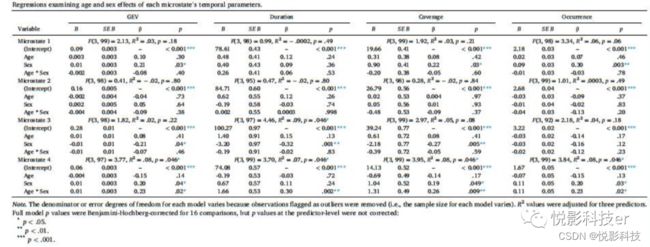
所有预测因子对微状态1和2的时间参数影响不显著。所有预测因子解释了观察到的微观状态3持续时间变化的显著比例,性别的主效应显著的,男性的微观状态3持续时间高于女性。所有预测因子对微观状态4的所有时间参数都有显著预测作用,简单分析结果如图3所示。
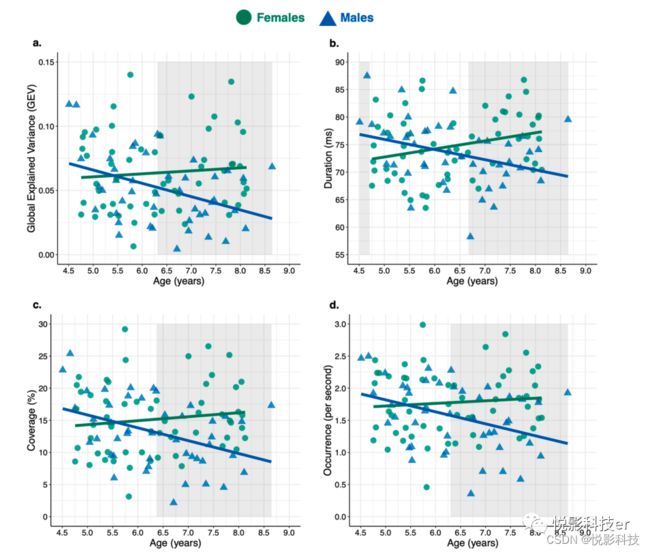
图3.年龄和性别对微观状态4的时间参数的交互作用。阴影区域代表具有显著性的 Johnson-Neyman区域(即,在p<.05时,性别的简单斜率显著不同于0的年龄区间)。
3.3 微观状态的溯源
四种微观状态的溯源估计和阈值为所有被试中激活的第95百分位以上的位点。从每个源图中减去四个阈值源图的平均值。考虑到年龄和性别对微观状态3和4的时间参数上的影响,将它们的来源叠加在图4的MRI上。
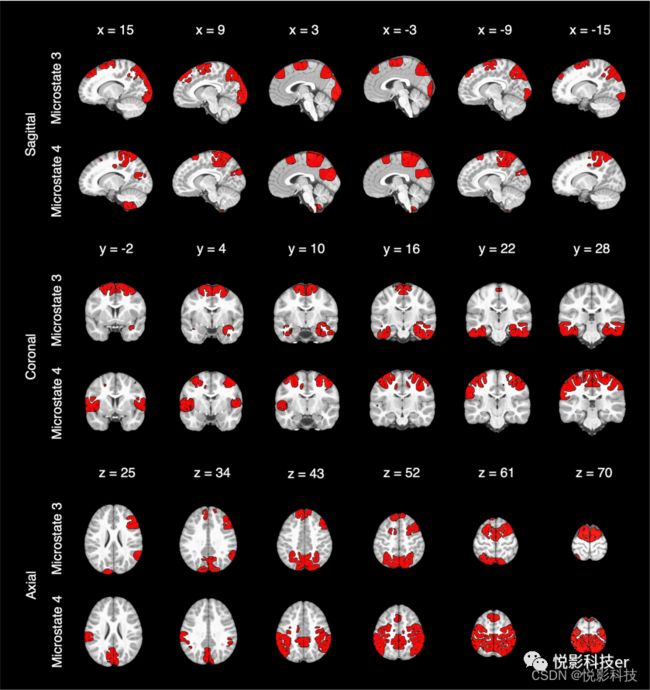
图4.微观状态3和微观状态4的溯源结果
4.讨论
与Tomescu等人(2018)一致,微观状态3和状态4的时间参数与年龄和性别有关,而微观状态1(典型A)和状态2(典型B)的时间参数与年龄和性别没有关系。微状态3持续时间男性与女性相比更为突出,微观状态4的所有时间参数都显示了年龄和性别的交互作用。男性的微观状态4 的GEV、覆盖率和发生率均随年龄的增加而降低,而女性则不随年龄的增加而变化。男性微观状态4持续时间随年龄而减少,女性随年龄而增加。事后分析显示,在所有微观状态4模型中,性别之间的年龄斜率差异在大约6.5岁时从不显著变为显著。此外,脑电图源定位表明,观察到的效应可能与注意和认知控制相关的网络有关。
本研究虽然是横断面研究,但也是第一次将6岁以下儿童典型的静息状态脑电图用于微状态分析。通过研究4-8岁儿童的大样本,获得足够的年龄变化来评估微观状态参数和年龄之间的连续关系以及他们与性别的相互作用,而不依赖按特定年龄分组的个体之间的群体水平统计分析。此外,本研究也是第一个评估儿童微观状态的潜在神经来源的研究,为脑电图微观状态在研究功能性大脑功能发育中的应用提供了希望。但研究是探索性的,而不是理论驱动的,所以结果应该谨慎解释。
参考文献:Spatiotemporal dynamics of EEG microstates in four- to eight-year-old children: Age- and sex-related effects