《Protein Actions Principles and Modeling》-《蛋白质作用原理和建模》中文分享(5)
《Protein Actions Principles and Modeling》-《蛋白质作用原理和建模》
本人能力有限,如果错误欢迎批评指正。
第二章:Proteins perform cellular functions
(蛋白质的细胞功能)
在一个细胞里合成一个全新的生物分子需要多步化学反应。这些化学反应的位点在空间定位上若是十分相近,那么会很大地提升合成的效率。有时候,一个酶分子就在空间中存在多个化学位点,这些位点分别存在不同的反应。而酶催化的过程中,这些不同的位点会分别负责不同的化学反应步骤。例如,脂肪酸合成酶,一种将两个碳添加到脂链中的二聚体酶,在其不同的结构域上催化一系列反应(图2.6)。动物的脂肪酸合成酶是二聚体,每个单体都具有脂肪酸合成所需的所有的酶结构域(每个颜色不同并在图中标记)。

图2.6 催化剂在空间的相邻性有助于生物化学反应的效率。脂肪酸合成酶是一种酶的局部复合物。白色箭头显示了脂肪酸合成中的五个连续反应,以将两个碳添加到碳氢化合物链中。所有反应发生在一个脂肪酸合成酶上。因为底物从一个结构域传递到下一个结构域:从丙二酰/乙酰转移酶(MAT,绿色)到酮酰基合成酶(KS,红色)到酮酰基还原酶(KR,黄色)到脱水酶(DH,蓝色)到烯酰基还原酶(ER,紫色)。灰色表示脂肪酸合酶的第二个相同单体。

图2.7 HIV逆转录酶(RT)将HIV的单链RNA复制到单链DNA上,以整合到宿主的基因组中。.RT具有两个变构偶联的催化位点:一个延长DNA,一个RNA酶H分解RNA。(底行):DNA聚合酶域的作用类似于拉动的手:手指(蓝色),手掌(绿色),拇指(红色),连接处(浅棕色)。RNase H“剪刀”域以紫色显示。结构支架以灰色显示。RT酶一次沿着病毒RNA移动一个碱基,将其复制到其补充DNA上,并通过一次去除一个核苷酸来切割原始链。(A)空酶。(B)聚合酶位点附近的结合多核苷酸链(黄色),具有由原始多核苷酸和新合成的链形成的双螺旋。(C)聚合酶的封闭形式。然后,链通过结构中的中央铰链运动向前拉动。
DNA和RNA聚合酶是添加核苷酸以生长DNA或RNA链的酶。RNA聚合酶将DNA序列转录成RNA序列。逆转录酶执行反向操作,复制RNA分子以产生DNA分子(图2.7)。例如,HIV逆转录酶(RT)具有两个配位的活性位点:一个位点用于复制病毒RNA,以及另一个在蛋白质的RNase H域上的位点,从复制的RNA链切割一个核苷酸。解旋酶是能量驱动的运动蛋白。它沿着双链DNA或RNA的链拉动自己,缠绕或解开旋转链,从而接近读取或修复核酸链的蛋白质。图2.8显示了拓扑异构酶的作用,拓扑异构酶是控制DNA双螺旋结构缠绕的蛋白质,这是为了获得复制DNA的蛋白质而需要的过程。

图2.8 拓扑异构酶II起到“通过另一个双螺旋的DNA双螺旋”的作用。(a)酵母拓扑异构酶II的结构。(B)(A)DNA双螺旋编号1进入蛋白质顶部,而DNA螺旋编号2已经绑定。(b)ATP水解后,螺旋2被切割。(c)螺旋1通过螺旋2中的断裂。(d)经过螺旋2后,DNA螺旋1现在从蛋白质中释放,并且螺旋2中的断裂被重新连接。
步行是将能量转换为运动的方法之一。你抬起你的左后腿,向前摆动,放下,抬起你的右后腿,然后重复这个过程。图2.9显示了的蛋白质的步行过程。驱动蛋白质有两只“脚“的蛋白。每当ATP分子与驱动蛋白结合时,驱动蛋白的后脚就会从轨道中拔出,向前走,然后在另一只脚的前边重新与轨道结合。在这种情况下,分子世界的行为模式就像它在腿和脚的宏观世界中一样。处于历史原因,驱动蛋白的脚被称为“头”。为了让蛋白质行走,它需要一条轨道来赋予它牵引力和方向。
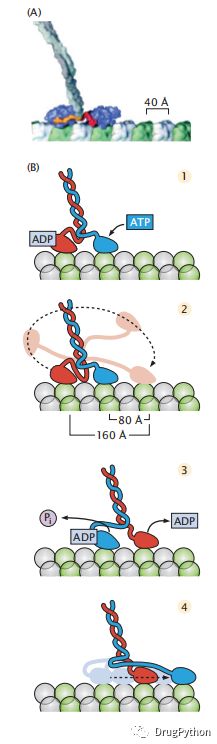
图2.9 驱动蛋白是一种运动蛋白,通过沿着微管轨道行走来携带东西。(a)驱动蛋白的两个头部的协调运动类似于行走。头部域(蓝色)与微管蛋白亚单元绑定,因为驱动蛋白沿着微管蛋白轨道(绿色/白色)行进。盘绕线圈(灰色)从轨道延伸到由驱动蛋白携带的货物。(B)该运动由ATP结合和水解在四个步骤的循环中驱动:(1)两个驱动蛋白头结构域都与微管蛋白结合。左边的也绑定ADP。当ATP与前边的头部结合时,运动启动,导致头部连接器(蓝色)与微管蛋白对接。(2)对接运动驱动另一个头部(红色)向前朝其下一个微管蛋白结合位点移动。(3)然后将该前头部与微管结合,导致沿着细丝的80 Å的的距离位移。绑定有助于从前头部释放ADP。同时,附着在后头部上的ATP被水解,释放磷酸盐(Pi)。(4)在ATP绑定到前头部之后,(蓝色)后头部向前移动。
轨道本身就是其他蛋白质分子(图2.10)。例如,驱动蛋白运动蛋白沿着称为微管的列车轨道状蛋白质四元复合物行进。微管是异二聚体蛋白组成的多股纤维。作为另一个例子,称为肌球蛋白的运动蛋白沿着肌动蛋白的轨迹行进。轨道原纤维通常由多个蛋白质链组成。作为另一个例子,称为肌球蛋白的运动蛋白沿着肌动蛋白的轨迹行进。轨道原纤维通常由多个蛋白质链组成。为了避免交通堵塞,根据细胞中的条件,这些轨道的微管蛋白亚单元在需要时组装和拆卸。从隐喻的角度来看,这就像是无论何时何地需要它们都会出现和消失的道路。微管蛋白丝的生长和收缩由游离微管蛋白二聚体的浓度和GTP的结合和水解控制。该纤维具有正端,其中添加新的微管蛋白二聚体。而负端的二聚体在GTP水解成GDP后解离。因此,如果有充足的自由微管蛋白和GTP,轨道的增长速度将快于收缩。

图2.10 微管蛋白的结构及其微管轨道。(A)与GTP(红色)结合的微管蛋白异二聚体从负端到正端组装成(B)原丝。(C)原丝组装在刚性圆柱形微管中。
有些蛋白质则是旋转马达。它们就像是汽车发动机或者电子时钟一样旋转。与现实意义上的马达一样,分子马达也有自己稳定的外壳。这些稳定部分常常嵌入在脂双层膜中。而旋转马达的灵活内部零件(如转子)都依赖于蛋白质提供的能量以特定的方向旋转。图2.11展示了称为F0F1 ATP合成酶的分子旋转马达,也称为ATP酶。细胞膜内外的PH梯度差异导致了质子流,从而ATP酶的转子转动,利用ADP和磷酸盐合成ATP。
在现实世界中,有些机器运作是可逆的。例如,应用电气电压驱动马达的转动,但是,相反,如果转动马达的轴,它会产生电压。有些蛋白质机器运作也是可逆的。ATP合成酶就是一个可逆的分子旋转马达。当ATP/ADP浓度的细胞质比高并且ATP水解反应(其释放能量)是有利的时,酶利用这种化学能泵送质子穿过膜以产生pH梯度。反之,当ATP低时质子向下流动,促使蛋白质从ADP合成ATP,补充细胞的ATP。详情见图2.11。

图2.11 F0F1 ATP酶是一种旋转运动蛋白,在质子梯度的驱动下合成ATP。(A)运动蛋白的侧视图。(B)细胞外部的pH值低于内部意味着外部的质子浓度较高。质子通过马达从外部(细胞外区域)向下流动到内部(细胞内)。这种质子流导致蛋白质旋转,进而导致蛋白质催化ADP转化为ATP。每个旋转周期合成三个ATP分子,对应于ATP酶环中的三个二聚体。
细胞膜是细胞内部与其周围环境之间的屏障。但是,如果细胞不能导入所需的营养分子或输出其废物,它就无法发挥作用。细胞必须进出口糖,离子,脂质,药物分子,有时还有蛋白质。细胞还必须通过选择性地渗透到各种离子和小分子来保持适当的平衡。这种运输对于神经元的电发射,肌肉细胞的收缩,卵子的受精,肾脏对小分子的过滤,细胞调节,中枢神经系统突触中信号的传递或神经递质的运输,以及食物的常规摄取和废物的输出是必不可少的。
膜蛋白是位于细胞内的脂质双层膜中的蛋白质。一些膜蛋白选择和调节分子进出细胞。对于某些膜蛋白,很容易从结构中解释功能。这些蛋白质具有穿过中心的孔,该孔充当穿过膜的管道。某些蛋白质可以更积极地控制转运。一些膜蛋白利用离子沿着膜上的电化学梯度(例如,从细胞外部到内部的钠离子)流动提供的能量来驱动其货物/底物(例如,神经递质如谷氨酸和多巴胺)的逆浓度梯度转运。这些蛋白质将离子的进膜流动与底物的出膜转运结合在一起。因为他们的运动是需要阳离子的协调转运才能实现,所以这种蛋白质被称为二级载体。相反的,只需要ATP水解获得能量的蛋白质被称为初级载体。
离子的转运过程通常需要满足各种要求。1.选择性:只有特定的离子可以进来或者出去2.方向性:离子流向一个方向,3调节性:离子的通量可以通过细胞膜的电压差,pH值,温度,外力或压力或与配体结合进行调节。离子泵根据浓度梯度主动输送离子。离子通道中的离子则是选择性地被动扩散到低电化学梯度的环境。例如,钾通道允许钾离子沿细胞膜的电化学梯度流动(图2.12)它们的选择性过滤器是氨基酸的空间排列,充当一种化学“筛”,通过钾并阻挡其他离子,甚至是其他小的正电荷离子。
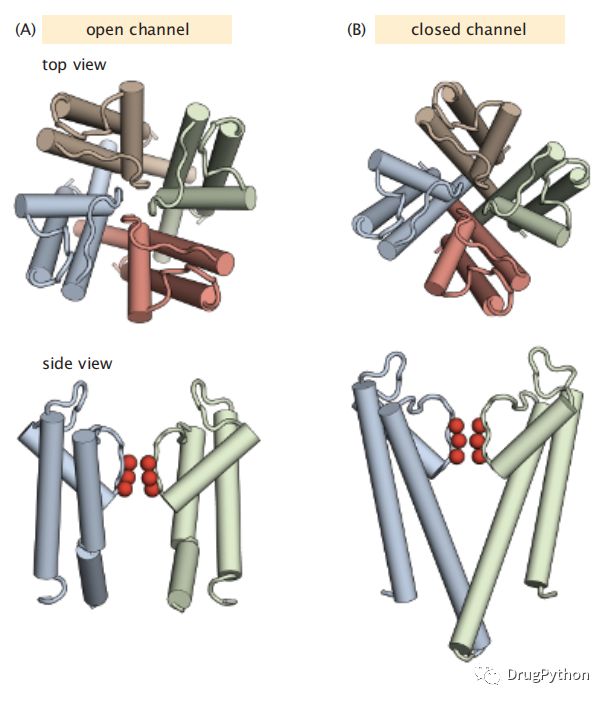
图2.12 KcsA是一个钾通道,其中心打开和关闭,允许钾流入细胞。KcsA是一种四聚体(每个亚单元在这里以不同的颜色显示),仅将钾离子从细胞的外部传导到内部。KcsA的跨膜螺旋显示在开口(A)和闭合(B)。最上面的一行显示了从顶部向下看通道时看到的四聚体的打开和关闭状态。下一行显示了仅两个单体的侧视图,其中羰基(红色)充当“选择性过滤器”以允许K+离子通过并阻挡其他离子。
有些膜通道蛋白是机械传感器:它们感知机械力并相应地增加或减少膜上的离子流。如果双层膜通过施加外力或压力在其平面中拉伸,则它可以导致膜通道蛋白的打开或关闭,导致离子在膜上的流动发生变化。MscL(大电导机械敏感通道)或MscS(小电导机械敏感通道)就是很好的机械传感器膜蛋白例子。在各种组织和生物体中都有机械敏感离子通道。他们在触摸,听力,平衡和痛苦中发挥作用。当细胞内的盐浓度变得太高,膨胀细胞,拉伸膜,并导致机械传感器通道打开离子通量并降低渗透压时,机械传感器蛋白还充当渗透压“释放阀”。
细胞内外有数千种不同类型的分子,从小到大,从带电到非极性。分子进出细胞必须是选择性的-细胞需要保留某些分子并排出其他分子。此外,并不是所有运输的“货物”分子细胞都是见过的(例如细胞以前没有见过的毒素)。那么这种高度特异性分子过滤器必须是具有选择性的。对于每种类型的东西都不可能有不同类型的受体。ABC转运蛋白是将不同类型的好分子导入细胞内部,而不会为坏分子打开通道。这些蛋白质允许不同的分子,如糖,维生素或金属。ABC转运体还出口疏水分子,如脂质,毒素或药物进入细胞内。另外,在致病菌中,一些药物虽然可以进入其细胞膜并杀死它们。但是这些细菌的ABC蛋白载体可以排出药物,会限制其有效性。

图2.13 ABC转运蛋白是膜蛋白,可以在细胞膜上运输许多不同类型的小分子。(A)ABC运输车的侧视图(绿色)。这种转运蛋白将维生素B12(红色)导入细胞。(B)这种运输蛋白具有多样性的。它可以与各种不同的对接模块蛋白(棕色)结合。每个对接模块本身识别并绑定到特定的小分子以导入或导出它们。一旦对接,小分子通过主动ATP驱动过程插入载体蛋白。使用ATP水解的能量,从而使载体蛋白的构象变化以打开孔来释放细胞上的配体,并且释放小分子进入细胞膜。
图2.13显示了大肠杆菌的维生素B12转运蛋白的工作原理。输送过程有两步。首先,BtuF蛋白是一种独立的“移动载体”,在细胞外捕获和结合维生素B12。BtuF将该物质运送到细胞膜,在那里它附着在对接模块膜蛋白BtuCD上。对接后,BtuCD将维生素B12穿梭穿过膜并将其释放到细胞内。该过程由提供能量源的核苷酸结合域提供动力。
-------------------------------------------
欢迎点赞收藏转发!
下次见!
