23《Protein Actions Principles and Modeling》-《蛋白质作用原理和建模》中文分享
《Protein Actions Principles and Modeling》-《蛋白质作用原理和建模》
本人能力有限,如果错误欢迎批评指正。
第五章:Folding and Aggregation Are Cooperative Transitions
(折叠和聚合是同时进行的)
-蛋白质可以协同组装成聚集体、原纤维或晶体
现在,让我们开始了解另一种协同过程吧。蛋白质之间可以互相聚集形成多蛋白聚合体(multiprotein assemblies),这个过程会随着浓度的变化进行急剧的变化(图 5.12)。该过程会导致蛋白质复合物或聚合体的形成,或聚集、沉淀、结晶、淀粉样蛋白生成或纤维化。例如,蛋白质可以结晶成天然分子相邻排列的对称重复形态。结晶很重要,因为它可以通过x射线衍射或散射来确定蛋白质的结构。多蛋白聚合体也发生在包涵体(inclusion bodies)中:某些生物技术会对生物体进行设计,使其过度生产一种感兴趣的特定蛋白质。过量生产导致蛋白质以高浓度积累,这些蛋白经常聚集成被称为包涵体的结构,该结构实际上包含了错误折叠或天然蛋白质的大量集合。在我们了解纤维化的特定模型之前,让我们来看看蛋白质聚集的一般性质。
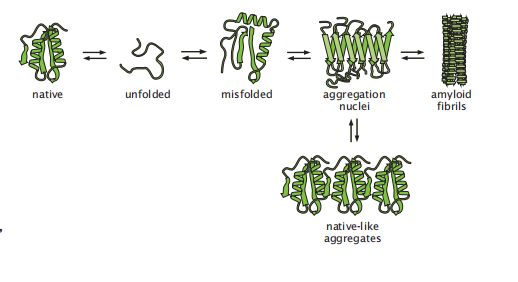
图5.12 蛋白质可以具有不同的状态,例如天然、展开、错误折叠、聚集、结晶或纤维。
吸引力的相互作用可以导致蛋白质聚集
蛋白质聚集的具体过程现在还不是很清楚,有些过程是不可逆的,因此无法使用平衡理论进行研究。但是有些规律依旧非常具有参考性。首先,带电荷的蛋白质会互相排斥。当蛋白质没有净电荷时,它们有最强的结合、聚集或结晶的倾向,当溶液的pH值等于蛋白质的等电点时,就会发生这种情况(图5.13 A)。第二,添加盐有利于促进具有大量净电荷的蛋白质的聚集。如果两个蛋白质分子具有相同的净电荷,添加盐可以屏蔽电荷,削弱斥力,促进蛋白质的聚集、纤维化和结晶等过程。例如,图5.13B显示,溶菌酶的溶解度随着氯化钠浓度的增加而降低,这意味着聚集性增加。第三,蛋白质分子可以通过疏水相互作用相互结合。聚集可以发生在具有疏水“粘性”补丁的蛋白质之间,或当内部疏水残基暴露时。例如,加热蛋白质通常会展开它们,至少部分地结构会有利于聚集状态。下面,我们将关注一种聚集类型的简单模型,即纤维形成,选择该模型是因为它是定量的,此外现有的实验给出了一些基本的见解。
淀粉样肽可以聚合(也可以称为组装)形成原纤维

图5.13 A蛋白质的溶解度取决于pH值和盐的浓度。(A)的溶解度在等电pH附近最小。红色曲线表示RNAse Sa的溶解度S(pI=为3.5);蓝色曲线表示pI =为6.4的RNAse Sa突变体的溶解度;棕色曲线表示pI =为10.2的突变体。需要注意的是,在野生型蛋白的情况下,最小值正好发生在pI = pH,在两个突变体中,由于其他效应(如偶极相互作用)它会轻微移动。(B)蛋白在高盐浓度下的溶解性较低。添加盐可以保护蛋白质上的净电荷,减少蛋白质之间的排斥,从而降低蛋白质的溶解度,促进聚集。这叫做盐析法。在其他情况下,通常不涉及带电蛋白质,增加盐可以增加溶解度,称为盐析。所示数据为溶菌酶。
有些蛋白质在浓度较高的时候会形成纤维状的聚合物。一根纤维就像是被包裹好的干意大利面:许多单独的链分子被相当直,排列,并紧密地排列在一起。纤维的浓度和蛋白质的浓度是相互关系具有一个非常大的变化。浓度低的时候,大部分的蛋白质处于游离状态并且以独立单体的形式在溶液中。但是,当蛋白质浓度达到某一个点的时候,这些蛋白质会急剧聚合,这种现象在多肽和蛋白质中都非常常见。原纤维和可溶性低聚物可在折叠性疾病中观察到,如阿尔茨海默氏病、帕金森氏病、亨廷顿氏病和朊病毒疾病。在某些情况下,一种具有正常稳定的天然结构的蛋白质可能会被扰乱,暴露出疏水表面,导致蛋白质粘在其他蛋白质上,形成聚集体。在其他情况下,那些没有稳定折叠结构的多肽也可以进行聚集。例如,Aβ是一种由40-42个氨基酸组成的肽,与阿尔茨海默病有关。α-突触核蛋白有140个氨基酸,具有一个高电荷和非结构的44个残基c端,与帕金森病有关。在亨廷顿氏病中,亨丁氨酸n端延伸的谷氨酰胺序列(例如,超过40个)可以导致聚集到斑块中。

图5.14 纤维的形成可以分为两个步骤:成核和增殖。(A)在成核过程中,前两条链聚在一起,开始形成原纤维。(B)在增殖过程中,额外的链增加到生长的纤维中。
图 5.14 展示了一个单体-纤维平衡的简单协同模型。在这里,我们主要计算不同大小纤维的浓度。设[A1]表示溶液中单体链的浓度(以单位体积中原纤维的数量为单位)。[A2]是含有两个蛋白质链的原纤维的浓度,[Am]是含有m个蛋白质链的原纤维的浓度。让我们将我们的自变量表示为x = [A1],以简化使用绑定多项式的标准符号(见第4章)。我们现在用孤立链的浓度x来表示[Am],如下。首先,将具有m跟链的纤维的浓度表示为将一条链加入m-1链个数的纤维的结合平衡:

其中K是将一个蛋白质添加到生长的纤维中的结合常数。重新排列方程5.24给出:
![]()
公式5.25描述了纤维的增殖过程,从m−1的链个数生长到m链个数的纤维;见图5.14B。
也就是说,每一条增加纤维的链乘以Kx。此外,根据螺旋-线团理论的相似想法,我们也定义了一个纤维成核过程。让成核平衡量δ解释第二个蛋白加入第一个蛋白在原纤维时开始形成:
![]()
这里,(Kx)是传播参数,δ是成核参数(类似于螺旋-线团理论中的s和σ)。δ<<1意味着启动原纤维的形成是困难的。纤维化模型和螺旋-线团理论之间的区别在于,纤维的形成取决于蛋白质的浓度,而螺旋-线团理论,它只描述了单个孤立分子的构象,并没有涉及到蛋白质的浓度。我们在这里建立的协同过程是一个结合平衡的问题,而不是构象平衡。
为了获得游离蛋白质浓度为x时的[Am]的浓度,我们需要乘以(Kx)m−1因为该模型通过增加m−1链上进行增殖,此外,我们需要乘以δ来表明启动了纤维进入二聚体。所以:
![]()
如果知道x的值,可以使用公式5.27来计算所有物种的相对浓度。但是,在进一步研究之前,让我们改用另一种表达相对浓度的常见方法。我们上边的计算[Am]指的是含有m链的纤维在单位体积的数量。而现在,我们想要计算蛋白质而非纤维。我们使用cm来表示含有m链的纤维上的蛋白质分子在单位体积的数量。即:cm=(每原纤维m蛋白)×(原纤维浓度):cm = m[Am] =(δ/K)m(Kx)m。c1为游离蛋白分子的浓度,c2为2-二分子原纤维中蛋白质分子的浓度,以此类推。那么溶液中蛋白质的浓度则可以如下所示:

你可以看到,方程5.28类似于方程5.14中的螺旋-线团配分函数。两者都是所有可能状态的统计权重之和。两者都有一个成核平衡常数(σ用于螺旋-线团过程,δ用于单体-纤维聚合)。两者都有一个增殖常数(s用于螺旋-线团过程,Kx用于单体-纤维聚合)。如果成核过程是不同的,那么单体-纤维的聚合和螺旋-线团的转变都是双态的。在这两个过程中,都不存在中间状态。对于螺旋-线团过程,如果σ很小,那么链将是一个线圈或一个长螺旋,而不是一个短螺旋。对于单体-纤维过程,如果δ很小(成核困难),那么大多数蛋白质要么以单体的形式,要么在大原纤维中,而不是在小原纤维中。支配单体和原纤维之间平衡的一个关键因素是Kx:K较大或蛋白质浓度x较大,那么整个环境主要为纤维状态。溶液中蛋白质浓度的增加会导致纤维状态的产生。
以下是该模型的三个主要结论: (1)对于给定的蛋白质浓度,含有m个链的纤维的分布[Am]呈指数级下降(图5.15A)。(2)在溶液中加入更多的蛋白质会导致原纤维的链的数量协同增加(见图5.15B)。(3)在溶液中加入更多的蛋白质会增加纤维的浓度,且呈非线性关系(见图5.15C)。
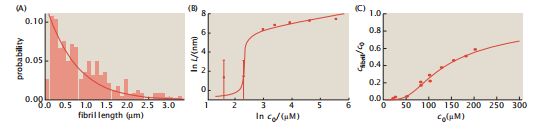
图5.15 淀粉样原纤维的生长。(A)淀粉样蛋白中原纤维的长度分布。条形图显示了淀粉样肽α-突触核素的实验粗度分布。(B)淀粉样原纤维随着肽浓度的变化而长得更粗。(C)原纤维的浓度随α-突触核蛋白的浓度呈非线性增加。
协同性是生物机制的一个共同特征。例如,病毒衣壳似乎一次从许多蛋白质分子中以全部或全部的方式组装起来。此外,当多个马达分子协同工作时,分子马达的力和速度——即沿着蛋白质轨道移动以进行蛋白质运输或产生力和流动的力和速度——会增强。在趋化过程中,细胞向食物移动的过程,受体蛋白在细胞膜上协同组装,以放大细胞对食物信号的检测。此外,蛋白质机器的组成部分聚合可能是协同的,但目前知之甚少。
-总结
我们已经了解了蛋白质中的各种协同过程。一些肽和蛋白质经历了一个尖锐的螺旋-线团转变,从一个大的变性构象集合到一个单一的螺旋构象。经典的螺旋-线团模型涉及到成核和增殖。成核是比较困难的,但增殖稍微具有偏向性,所以链要么没有螺旋要么形成长螺旋。蛋白质的折叠一般也是协同的。二级结构和三级结构都有助于其稳定性和协同性。此外,我们还探讨了高浓度的蛋白质如何结合形成纤维聚合。
第五章完
-------------------------------------------
欢迎点赞收藏转发!
下次见!