百趣代谢组学分享-基于靶向PFKFB3阻断糖酵解抑制婴儿血管瘤的发展

代谢组学文章标题:Blockage of glycolysis by targeting PFKFB3 suppresses the development of infantile hemangioma
发表期刊:Journal of Translational Medicine
影响因子:8.44
作者单位:四川大学华西医院
百趣提供服务:中心碳代谢
代谢组学分享-研究背景
婴儿血管瘤(Infantile Hemangioma, IH)是婴儿最常见的肿瘤,但其确切的发病机制尚不清楚。前期研究发现,糖代谢可能在IH发病机制中发挥重要作用,抑制糖酵解关键酶磷酸果糖激酶-1可抑制IH血管生成。代谢组学分享,磷酸果糖-2-激酶/果糖-2,6-双磷酸酶3 (PFKFB3)是一种将果糖- 6-二磷酸转化为果糖-2,6-二磷酸的代谢酶,是限速酶磷酸果糖激酶-1最有效的变构激活剂。本研究旨在探讨PFKFB3在IH中的作用。
代谢组学分享-结果与分析
1.PFKFB3在增殖期IH组织与HemEC中高表达
共鉴定出222个差异表达基因(Differentially Expressed Genes, DEGs)(141个上调,81个下调),如下图所示(图1A),PFKFB3是最显著的DEGs之一。DEGs在生物过程中主要富集在 “细胞脂代谢”等过程中(图1B);在KEGG途径中主要富集在“甘油脂代谢”等通路上(图1C)。代谢组学分享,在对芯片结果验证过程中,发现PFKFB3蛋白在增殖型IH组织中的表达高于渐开化型消退期IH组织(图1D)。此外,PFKFB3蛋白在血管瘤内皮细胞(Hemangloma-derived endothelial cell, HemEC)中的表达高于人脐静脉内皮细胞(Human umbilical vein endothelial cells, HUVECs)(图1E)。
总之,细胞代谢与IH的发展密切相关,PFKFB3在增殖的IH组织和HemECs中高表达,可能在IH发展的调节中发挥重要作用。

图1. PFKFB3在增殖IH组织和HemEC中的过表达
2.PFK15抑制HemEC血管生成
随着药物浓度的增加,PFK15(PFKFB3抑制剂)处理引起HemECs的形态学逐渐改变,包括细胞密度下降、细胞收缩和细胞碎片增加(图2A),PFK15以剂量依赖的方式降低HemECs的活力(图B)。代谢组学分享,为了进一步验证PFK15在抑制细胞增殖中的作用,在体外评估了PFK15对HemEC体外成管的影响。结果表明,PFK15显著抑制血管生成(图C)。总之,用 PFK15抑制PFKFB3可以降低HemEC血管生成。

图2. PFK15抑制HemEC血管生成
3.PFK15抑制HemEC葡萄糖代谢
对葡萄糖进行靶向代谢组学分析。PCA(图3A)图显示对照组和PFK15处理组之间存在显著差。如火山图(图3B)和热图所示(图3C),共鉴定出45种差异代谢物。PFKFB3被抑制后,许多糖酵解代谢物,包括葡萄糖6-磷酸、果糖6-磷酸和果糖1,6-二磷酸含量仍比对照组更高(图3C)。与对照组相比,PFKFB3被抑制后,属于“TCA循环”的代谢物柠檬酸、异柠檬酸、富马酸、α酮戊二酸、苹果酸和琥珀酸也显著增加(图C)。代谢组学分享,随后选取差异最高的14种代谢物进行KEGG通路分析。发现这些代谢物主要富集在糖酵解相关途径,如“磷酸戊糖途径”、“糖酵解/糖异生”、“丙酮酸代谢”和“TCA循环”(图3D,E)。综上所述,被PFK15抑制的PFKFB3可以减少HemECs的糖代谢。

图3. PFK15抑制HemEC葡萄糖代谢
4.PFK15抑制HemEC迁移,诱导细胞凋亡
细胞迁移实验结果显示PFK15抑制HemECs的迁移(图4A)。此外,当PFK15联合普萘洛尔(propranolol)治疗时,观察到一种协同抑制作用(图4A)。代谢组学分享,为了确定PFK15诱导的协同迁移抑制是否是由于细胞凋亡,使用Annexin V和PI标记进行流式细胞术分析。结果显示,PFK15处理组的凋亡细胞数量大于对照组(图4B)。
同样,PFK15和普萘洛尔联合治疗导致HemECs凋亡百分比显著增加(图4B)。此外,通过western blot分析,PFK15和HemECs联合治疗后Bax蛋白表达上调,Bcl-2蛋白表达下调(图4C)。代谢组学分享,与对照组相比,PFK15处理后观察到明显的线粒体应激反应,包括线粒体基质密度增加,线粒体嵴塌陷,线粒体肿胀和线粒体空泡变性(图4D)。线粒体应激反应可大大增加活性氧的产生,导致线粒体凋亡通路的激活。结果显示,与对照组和普萘洛尔组相比,PFK15治疗后ROS生成增加(图4E,F)。
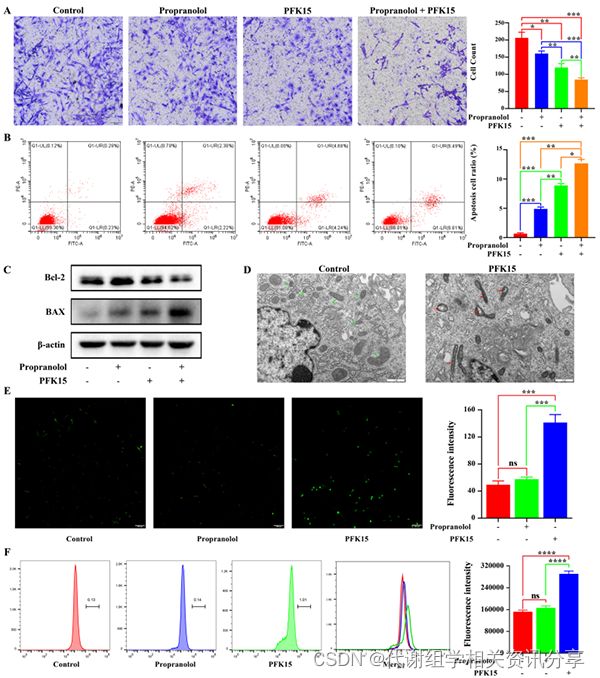
图4. PFK15抑制HemEC迁移并诱导细胞凋亡
5.shPFKFB3抑制HemEC血管生成
为了进一步研究PFKFB3水平是否影响血管生成活性,我们构建了靶向PFKFB3的慢病毒载体shRNA(图5A)。代谢组学分享,转染shPFKFB3后,PFKFB3在HemECs中的蛋白表达水平显著降低(图5B)。
敲低PFKFB3也显著抑制了HemEC的血管生成(图5C)。shPFKFB3显著抑制了细胞迁移(图5D)。此外,shPFKFB3慢病毒组的凋亡细胞比例高于对照组(图5E)。综上所述,敲除PFKFB3显著抑制了HemEC血管生成。
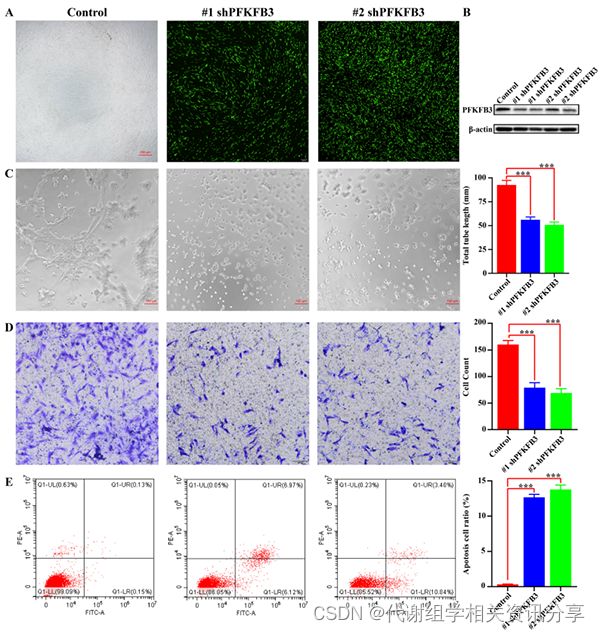
图5. PFKFB3下调抑制HemEC血管生成并诱导细胞凋亡
6.PFKFB3抑制降低了HemECs中PI3K/Akt信号通路的激活
为了确定PFKFB3被抑制后IH中调控血管生成活性的关键通路,对PFK15处理组和对照组进行了转录分析。共鉴定出1240个DEGs(357个上调,783个下调)。GO分析显示,这些DEGs主要富集在“血管生成”、“细胞粘附”和“凋亡过程的正向调控”等生物过程中(图6A)。KEGG通路分析中显著富集在PI3K-Akt信号通路(图6B)。这些数据表明,PI3K-Akt通路可能参与PFK15治疗后IH血管生成和细胞代谢的调节。代谢组学分享,此外,对转录和代谢结果进行综合分析的结果显示,“糖酵解或糖异生”是最重要的途径(图6C),表明抑制PFKFB3主要影响HemECs中的糖酵解。
为了进一步证明PI3K-Akt通路在PFKFB3调控中的作用,我们首先使用STRING进行了功能蛋白关联分析,以建立Akt和PFKFB3之间的潜在关系。可以看出,PFKFB与Akt直接相连,Akt也与Bax和Bcl-2密切相关(图6D)。接下来,利用Co-IP验证PFKFB3与Akt之间的蛋白-蛋白相互作用(图6E)。
由于PI3K/Akt信号通路参与了PFKFB3的调控,我们试图确定PFK15是否通过该通路诱导HemEC凋亡。Western Blot结果显示,PFK15降低了PI3K和Akt的磷酸化水平(图6F)。代谢组学分享,与单独使用每种药物相比,PFK15和普萘洛尔联合使用可进一步降低PI3K/Akt的磷酸化水平(图6G)。此外,用shPFKFB3敲除PFKFB3可显著降低PI3K和Akt的磷酸化水平。因此,通过抑制PFKFB3可抑制PI3K-Akt信号通路,诱导细胞凋亡。

图6. PFKFB3抑制抑制HemECs中PI3K/Akt信号通路的激活
7.抑制PFKFB3可抑制体内IH血管的形成
为了研究我们的体外研究结果是否可以转化为体内环境,我们使用HemECs和血管瘤源性周细胞建立了小鼠异种移植模型(图7A)。14d处死实验动物,解剖实验动物的肿瘤(图7B)。代谢组学分享,14d后仅PFK15组和联合用药组的肿瘤生长显著降低(图7C)。
HE染色结果显示,PFK15单独或联合普萘洛尔可显著减少微血管数量(图7C)。此外,与对照组相比,PFK15单独使用或联合普萘洛尔可降低CD31基因的表达(图7C)。转导shPFKFB3可减少微血管数量和CD31的表达(图7D),沉默PFKFB3也显著降低了MVD(图7F)。所有这些结果都表明,抑制PFKFB3可以抑制体内IH血管的形成。

图7. 抑制PFKFB3可抑制体内IH血管的形成
代谢组学分享-结论
总之,这些结果表明抑制PFKFB3可以降低IH血管生成并诱导细胞凋亡,靶向PFKFB3可能是IH的一种新的治疗策略。