病原菌共感染研究思路

近年来,多微生物共感染在临床上的报道日益增多。其中,多细菌共感染占据了细菌感染的25% ,其发病率和严重性也相应增加。尤其在形成生物膜后,这种共感染对疾病的发生、发展和临床治疗生重要影响,同时也给临床治疗带来了更大的挑战。
随着测序的发展,应用高通量转录组测序对于感染多种病原菌的研究也不断深入,今天跟大家分享两篇病原菌共培养研究共感染过程的相互作用,希望能给研究共感染的老师一些样本准备设计的思路的参考。
案 例 一

在囊性纤维化患者中常出现铜绿假单胞菌和金黄色葡萄球菌的共同定植,且患者预后较差,然而病原体之间的互作研究较少。首先跟大家分享一篇来自BMC Genomics 的名为Transcriptional profiling of Pseudomonas aeruginosa and Staphylococcus aureus during in vitro co-culture文章,作者通过对铜绿假单胞菌和金黄色葡萄球菌共培养不同时间点进行转录组测序,分析共培养过程中两种菌株的相互作用关系。
-
研究背景
微生物在共享同一环境时,可以进行互惠、中性或拮抗的相互作用。在感染过程中,细菌会遇到营养不良的环境,导致资源竞争和抗生素作用。细菌之间的合作也会发生,铜绿假单胞菌(P. aeruginosa)通过形成混合生物膜保护金黄色葡萄球菌(S. aureus)免受盘状盘基网柄菌的吞噬。
在本研究中,作者将铜绿假单胞菌和金黄色葡萄球菌单菌培养转录组以及共孵育3小时后的转录组比较分析。数据结果展示出两种细菌的基因表达变化主要包括核苷酸和氮代谢、毒力因子和噬菌体相关基因的诱导表达。
-
技术路线

图1 共培养技术路线图
-
结果展示
共培养时两种菌CFU比值约为50%与t0时初始比值相当,说明一开始的竞争并没有马上影响各自种群变化。4h金黄色葡萄球菌单独培养CFU和共培养菌体CFU都开始下降,并且变化趋势一致。
通过转录组测序比较两种菌在共培养和单独培养状态下基因表达结果表明,每种细菌的基因表达都受到另一个菌存在的影响。与单培养相比,铜绿假单胞菌共培养时基因表达下调和上调的基因分别为1726和1718,金黄色葡萄球菌共培养与单培养相比基因表达下调761和上调的基因为730。表达差异超过2倍((log2 ratio ≥1or≤-1)占铜绿假单胞菌基因组的9.4%(545个基因)和金黄色葡萄球菌基因组的18.6%(450个基因)。其中,铜绿假单胞菌中上调的基因有181个(33.2%),下调的基因有364个(66.8%);金黄色葡萄球菌上调的基因有189个(42.1%),下调的基因有261个(47.9%)(表1,图2)。差异基因功能注释表明,代谢和转运是影响最大的分类。有趣的是,这些类别的比例在这两种生物中是相似的,代谢功能为 28.8% (P. aeruginosa)和 27.8%(S. aureus),转运功能为12.5% (P. aeruginosa)和13.3%( S. aureus)(图3)。RNA-seq数据通过实时荧光定量 PCR对每个细菌的6个基因(3个上调和3个下调基因)进行了验证。


图2 共培养与单培养基因功能分类变化
-
结 论
对金黄色葡萄球菌和铜绿假单胞菌共培养进行转录组分析显示,在这些细菌病原体初次接触期间,它们之间存在复杂的适应模式。差异表达基因很大部分属于功能类“代谢”和“运输”。很有意思的是,铜绿假单胞菌驱动金黄色葡萄球菌进入发酵,前者利用后者产生的乳酸作为碳源。这两种生物都表现出氮饥饿的迹象,可能诱导金黄色葡萄球菌的嘌呤和嘧啶合成途径。在其中并没有发现铜绿假单胞菌诱导抗葡萄球菌通路的基因表达,这表明在共培养的早期阶段,主要是资源竞争而不是直接(干扰)竞争,这两种生物似乎都能感觉到营养剥夺、ROS介导的攻击或直接细胞损伤造成的压力。
案 例 二
瘤胃中微生物群落关系复杂,对瘤胃真菌及其与其他微生物群落成员之间的相互作用研究较少。瘤胃真菌(厌氧肠道真菌)在大型食草动物的消化道中占有相当重要的作用,其与细菌、产甲烷古菌和原生动物作为生物降解的主要组成部分。瘤胃中细菌的数量至少比真菌多4个数量级,但两者的相互作用也一直是广受关注,来自美国加州大学Candice L. Swift发表的题为Cocultivation of Anaerobic Fungi with Rumen Bacteria Establishes an Antagonistic Relationship文章利用转录组测序(RNA-seq)和代谢组分析了瘤胃细菌琥珀酸纤维杆菌(Fibrobacter succinogenes)菌株UWB7与厌氧肠道真菌 (Anaeromyces robust)或 (Caecomyces churrovis)共培养后各菌种基因表达以及和代谢物的变化,为后续新型真菌抗生素的筛选提供新的参考。

-
研究背景
瘤胃细菌和厌氧真菌是瘤胃微生物群落中的两个重要成员,它们在瘤胃中共同参与纤维素降解和产氢过程。然而,关于它们之间的相互作用和共生关系的了解还很有限。本研究通过对厌氧真菌和瘤胃细菌进行共培养实验,探究它们之间的相互作用,并研究了共培养对它们的代谢产物和基因表达的影响。这些研究有助于深入了解瘤胃微生物群落的功能和相互作用机制。
-
技术路线

图3 技术路线
-
结果展示
瘤胃真菌A. robustus/C. churrovis和UWB7共培养,直至真菌达到对数中期,此时将共培养和相应的真菌单培养进行RNA提取、建库(共培养物分成两份分别进行polyA捕获和rRNA去除两种建库方式)和测序。
无论是A. robustus 还是 C. churrovis与UWB7共培养培养都上调了编码应激反应蛋白的基因,这表明细菌的存在引发了真菌的应激反应,在不同碳源上都观察到真菌胁迫。差异基因富集分析显示共培养中上调的热休克蛋白富集在Avicel上共培养与在柳枝稷培养基上培养相比较,真菌共培养与单培养的差异基因数量大约是10倍。这种差异可能反映出菌株UWB7可以有效的利用植物纤维素,从而提高自己的菌株生长并对真菌产生影响(图4)。A. robustus与菌株UWB7共培养的条件下,与在Avicel培养基上生长单独培养的A. robustus相比,其次级代谢相关基因:6种非核糖体肽合成酶(NRPSs)和1种聚酮合成酶(PKS)样酶上调了至少2倍。A. robustus通过调节多酮和非核糖体肽的合成来应对细菌UWB7的攻击。

图4 在不同源培养基上A.robustus与菌株UWB7共培养基因表达热图
值得注意的是,A. robustus/C. churrovis与UWB7共培养均能上调s -腺苷-蛋氨酸(SAM)依赖的甲基转移酶、组蛋白甲基转移酶和乙酰转移酶。真菌组蛋白H3K27me3标记在共培养中表达增强,异染色质蛋白下调。总之,这些发现表明在细菌存在的时候真菌染色质重塑发生变化(图5)。
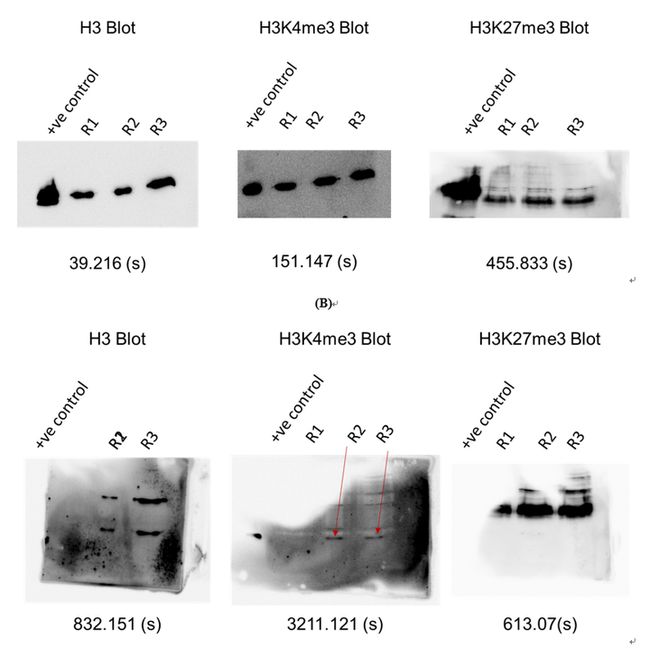
图5 真菌A.robustus组蛋白修饰变化WB
厌氧真菌与细菌的共培养表明有独特的代谢物分泌。为了进一步验证厌氧肠道真菌与琥UWB7共培养会触发真菌防御化合物的产生这一假设,对真菌-细菌共培养以及各自的真菌或细菌单培养进行了非靶向代谢组学分析。单培养中未发现m/z 244.227代谢物的特征,但在Avicel上共培养时,m/z 244.227的含量比单培养的A. robustus高12倍,表明这是一种独特的真菌代谢物,随着细菌UWB7的存在而增加(图6)。

图6 共培养状态代谢物分析
-
结 论
尽管之前的报道表明琥珀酸纤维杆菌与瘤胃真菌没有相互作用,此文中作者利用RNA-seq证明,通过共培养与单一培养相比较,菌株UWB7与A. robustus/C. churrovis共培养,会导致细菌和真菌转录组的急剧变化,包括细菌药物流出泵和真菌伴侣、聚酮o -甲基转移酶、PKSs和NRPSs的上调。此外,真菌基因编码的组蛋白修饰酶在共培养中上调。共培养组蛋白H3K27me3标记升高,异染色质相关蛋白下调。
总之,这些结果表明,类似于高级真菌,厌氧真菌通过染色质重塑调节其二级代谢。厌氧肠道真菌在受到瘤胃细菌挑战时通过表观遗传和转录调控激活其二级代谢。这些转录变化的代谢结果可能是由PKS或NRPS产生的真菌防御化合物的产生。与这一假设一致,非靶向非极性代谢组学发现至少一种独特的真菌代谢物通过共培养产生。因此,厌氧真菌和瘤胃微生物群的拮抗关系可能被证明是抗生素的一个宝贵的来源。

-
参考文献:
1 .Tognon M, Köhler T, Luscher A, van Delden C. Transcriptional profiling of Pseudomonas aeruginosa and Staphylococcus aureus during in vitro co-culture. BMC Genomics. 2019 Jan 10;20(1):30.
2 .Swift CL, Louie KB, Bowen BP, Hooker CA, Solomon KV, Singan V, Daum C, Pennacchio CP, Barry K, Shutthanandan V, Evans JE, Grigoriev IV, Northen TR, O'Malley MA. Cocultivation of Anaerobic Fungi with Rumen Bacteria Establishes an Antagonistic Relationship. mBio. 2021 Aug 31;12(4):e0144221.