Experimental evolution makes microbes more cooperative with their local host genotype
实验进化使微生物与其本地宿主基因型更合作
摘要:微生物学的发展需要更好地了解有益微生物如何适应宿主。为了探究固氮细菌(根瘤菌)如何适应豆科植物,作者进行了长达一年的进化实验和交叉接种实验,来测试宿主是否会选择更具协作性的微生物菌株。作者将敏感菌株Ensifer meliloti与五种苜蓿(Medicago truncatula)基因型分别进行进化实验,这些基因型苜蓿对共生细菌的选择强度不同。在不依赖宿主选择的情况下,EM能迅速适应宿主的基因型,而衍生(drive)的微生物在与宿主共享进化史时更有益。这种局部适应性主要局限于共生质粒可能发生的基因突变。因此,合作依赖于伴侣基因型之间的配对,并且随着细菌对该宿主的适应而增加。
宿主相关的微生物群通常是有益的,但我们对这些共生关系中伙伴之间的适应了解有限,尤其是在基因组水平上。互生有时被视为相互寄生,潜在地导致保持种群内遗传变异的对抗共同进化。然而,最近的研究质疑了互惠关系中适应冲突的普遍性,伴侣之间一致的适应性利益应该导致进化停滞,减少了伙伴质量等互惠性状的遗传变异。
劣质(poor-quality)微生物可能是“骗子”(cheaters),通过牺牲宿主为代价提高自己的适应性。然而,宿主经常“选择”他们的微生物群,并优先与更合作的微生物联系或给予奖励,选择那些潜在的骗子。这样选择是适应性过程,但是也是矛盾的:如果挑剔的寄主选择了高质量(high-quality)的共生体,那共生体的质量变化就会减少,减少了宿主对自身的选择优势。然而,很多宿主是挑剔的,许多共生体也不是非常的有益,那么这些无效(ineffective)微生物的存在就很令人困惑。
无效(ineffective)微生物可能只是与其宿主不匹配。一种宿主基因型上的高质量微生物可能在另一种宿主基因型上属于低质量微生物。这种共生体质量的基因型-基因型相互作用(GxG)很常见,维持了共生体提供给宿主的利益的变异,是共同进化的先决条件。局部适应,即来自同一位点的伙伴优于来自不同位点的伙伴,可以产生GxG相互作用并发生在互惠中,但它不是普遍存在的。此外,在局部适应或GxG相互作用的遗传机制仍然很大程度上是未知的。
豆科植物与根瘤菌的相互作用具有重要的经济和生态意义,是研究相互作用的一个模型。豆科植物通过根瘤菌将碳转化为氮,根瘤菌存在根瘤中。根瘤菌传代快,可培养,可通过水平基因转移获得突变。根瘤菌也适用于能够识别与表型相关的基因组变异的全基因组关联研究(GWASs)。Burghardt等人采用这些方法,发现寄主根瘤菌的选择性强于自由生活在土壤中的根瘤菌。
在这里,作者对根瘤菌在选择性不同的五种宿主基因型中的一种上进化后进行了重新测序和交叉接种,以测试更挑剔的宿主是否会选择更合作的共生体,或者根瘤菌是否会适应当地的宿主基因型。使用了两种不同宿主效益的根瘤菌菌株:不敏感的菌株Sm1021(这里简称为Em1021)和敏感的固氮菌菌株WSM1022 (Em1022)。然后将这两个株系与五种苜蓿自交系中的每种分别进行配对实验,270株系不加选择。而其他株系都更喜欢Em1022而不是Em1021,在过去的实验中,267株系几乎只与Em1022合作。经过历时一年、跨越5代植物的进化实验,作者从根瘤中分离出“衍生”根瘤菌,并将它们与原始的“祖先”(ancestral)菌株进行比较。
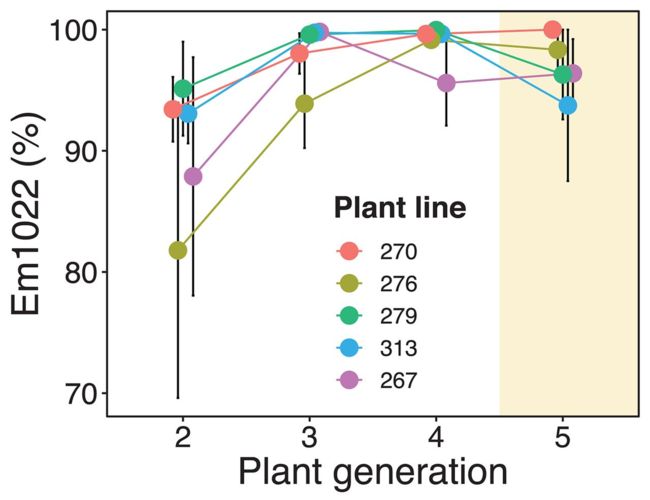
尽管Em1021开始时具有双重优势,但它几乎在所有寄主株系上灭绝(图1)。在实验结束时,Em1021只与15%的植株相关,在336个取样的结节中只占3%。在结合土壤和根瘤应变频率的线性模型中,发现世代的影响边际显著,而株系的影响不显著。有效的共生体,Em1022,在所有的寄主上超过了Em1021,而不管其选择性如何,这表明伴侣的选择并不是一个重要的选择力量。
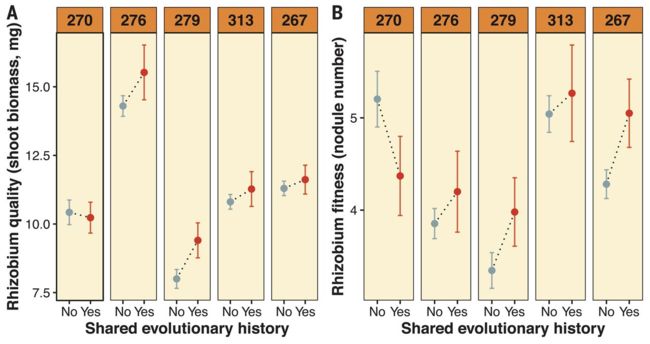
作者通过种植5个株系的新种子,分别接种原始的Em1022或40个衍生分离株中的1个,测试所有可能的宿主基因型和分离株组合的重复,来检测Em1022表型。作者通过测定地上部植物生物量量化根瘤菌对宿主的贡献,对40个衍生菌株的全基因组进行测序,并对每个祖先的9个重复进行测序,并对特定宿主基因型和所有株系的共生体质量和适应度进行GWASs检测。
实验进化的细菌给宿主带来的好处的差异很大程度上取决于根瘤菌和宿主基因型之间的匹配。衍生的Em1022和祖先的Em1022在共生体质量或适应度方面没有显著差异(表S1A和S2A)。然而,在进化实验中,衍生菌对与之配对的宿主基因型提供了更大的好处(图2A),尽管少数分离株是高或低质量的植物株系共生菌(图2A,S1和S2)。衍生根瘤菌在其本地宿主基因型上进行测试时也获得了更高的适应度,除了在最不挑剔的宿主基因型上进化的分离株(第270行)(图2B)。线性混合模型发现,共享的进化历史对根瘤菌质量和适应度都有显著的正向影响(表S1B和S2B)。
从所有宿主的平均值来看,根瘤菌质量和适合度之间存在正相关关系(图S3),这表明40个衍生菌株没有进化出欺骗,强烈的适合度反馈使合作适应根瘤菌。同一性状在不同宿主上的两两相关性较弱,通常在统计学上不显著(图S4),这表明一个宿主上的微生物质量或适应度很难预测另一个宿主上的质量或适应度。
苜蓿分离到的根瘤菌的基因组测序发现了1330个遗传变异,包括547个从头突变(图S5A)。多数从头变异(305)特异于最少选择宿主细胞系(第270行)产生的根瘤菌; 较少的变异体(16到59)是针对其他品系的根瘤菌特异的(图S5A)。在筛选出高连锁不平衡中的大多数单例和变体,突变基因减少到363个之后,又通过GWAS在一个或多个宿主环境(表)中鉴定了145个与根瘤菌质量或适应性显着相关的变体(称为重要突变体)。这些变体大多位于两个共生质粒pSymA(62个)和pSymB(68个)上(细菌染色体15个),它们分别含有根瘤菌生命周期的共生和自由生活阶段必不可少的基因。
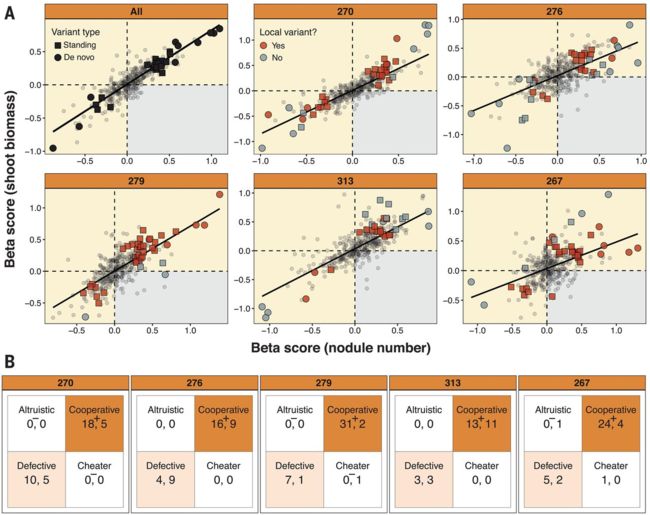
Beta值代表基因组变异与表型之间的关联强度,在共生体质量与适应性之间呈正相关意味着增加微生物适应性的基因组变异也增加了微生物对植物的益处。在每个宿主环境中,大多数重要变体在进化实验中与该宿主基因型配对的分离株中进化(即局部变体;图3A中的红色符号和图3B中的逗号前的数字)。如果共生体质量和适应性的β得分均大于零,则将变异效应分类为“合作”;如果共生体质量和适应性的β得分均小于零,则将变异效应分类为“缺陷”;如果共生体质量β值为正,但对宿主适应性的β值为负,则为“利他”;如果共生体质量β分为负,但对宿主适应性的β值为正,则为“骗子”。在所又5个宿主环境中,大多数显着的变异效果是协同的和局部的(图3B和表S3)。
通过将基因组随机分配给表型1000次并重新计算β得分,作者使用排列来评估这种模式是否可能是偶然发生的。在5%错误发现的情况下,所有五个宿主环境中的本地合作关系明显较多(图3B,加号),而在某些宿主环境中,本地利他主义和本地作弊者的关联(图3B,负号)也明显较少。其他类别(例如,有缺陷的变体)中的关联数量与零期望没有不同。
局部合作变体位于可能编码钙结合蛋白,黄素腺嘌呤二核苷酸(FAD)结合氧化还原酶和两种Ti型共轭转移松弛酶的基因中(表S3)。这些功能可能与结节内结节的发生或根瘤菌的存活有关(23)。变异效应通常是宿主特有的,并且通常对其本地宿主有条件的好处,或者对非本地宿主有害(表S3),这表明在本地环境中进行选择,在非本地环境中进行漂移。当变体对多个宿主表现出显着影响时,它们往往在宿主之间始终是有益或有害的; 只有两个变体在宿主之间具有显着的相反符号效应,这表明折衷很少见(表S3)。总之,由于细菌适应了其本地宿主基因型,因此偶然发生的合作比偶然发生的频繁,而欺骗或利他的发生则更少。
在进化实验中,作者期望最选择的宿主系最强烈地支持更好的共生体Em1022。然而,无效菌株Em1021在所有植物品系中几乎灭绝,甚至在没有选择性的寄主上也是如此(图1),这表明该菌株是普遍竞争较弱的菌株。相比之下,Em1022进化了以前未表征的变体,其共生体质量和适应性均不同。衍生细菌提供了更大的宿主益处,并且通常在共享进化史的宿主基因型上具有更高的适应性(图2),并且检测到显着过量的基因组变异对本地宿主具有协同作用(图3)。这些结果表明,与伙伴选择相比,局部适应是塑造微生物合作更重要的进化力量。此外,当微生物始终与同一宿主配对并且扩散受到限制时,所产生的局部适应性可能导致更多而不是更少的合作。出现了一种适应宿主-微生物共生的模型:微生物可以通过站立或从头变异迅速适应特定的宿主基因型,这些变异也有益于本地宿主,但对非本地宿主基因型有不同的影响,从而保持了共生质量的差异。观察与宿主相关的微生物。该结果还暗示了在密切相关的宿主之间移植微生物群更可能有效,或者需要给微生物足够的时间使其适应新的宿主环境。
原文地址:Experimental evolution makes microbes more cooperative with their local host genotype