项目文章 |首次发现太平洋亚历山大藻(甲藻)快速生长中H3K4me3修饰


发表单位:中国海洋大学海洋遗传育种教育部重点实验室
发表日期:2022年11月30日
期刊 :Frontiers in Marine Science (IF2022: 5.247)
2022年11月30日中国海洋大学海洋遗传育种教育部重点实验室在Frontiers in Marine Science 发表了题为“First insight into H3K4me3 modification in the rapid growth of Alexandrium pacificum (dinoflagellates)”的研究论文。该研究结果表明H3K4me3在调节太平洋亚历山大藻的快速生长中发挥着潜在的重要作用。这些关于这种甲藻中组蛋白修饰调控网络的知识,为未来相关领域的研究奠定了必要的基础。爱基百客为该研究提供ChIP-seq的技术支持。

1 研究背景
太平洋亚历山大藻是许多沿海地区主要的开花微藻物种之一。营养物浓度的变化,特别是氮的变化,与水华的发生和终止密切相关,氮的积累诱导藻类细胞的快速生长。然而,对太平洋亚历山大藻生长机制的研究,包括分子机制的表观遗传学调控,仍然有限。甲藻的基因组结构是独特的,非编码序列占基因组的60%,这表明存在着巨大而复杂的调控网络。因此,研究甲藻的这种调节作用对研究其生理活性和对环境的反应机制具有重要意义。组蛋白修饰作为表观遗传调控的一种形式,在植物生长、发育和应激反应中发挥着重要作用。然而,它们对藻类的影响仍然知之甚少,尤其是对甲藻的影响。
在本研究中,H3K4me3的表观基因组图谱被用于研究其在快速生长对高光和高氮的反应中的作用。本研究旨在通过ChIP-seq在太平洋亚历山大藻中产生第一个全局H3K4me3图谱,以揭示H3K4me3维持开花进程的调节机制,并破译快速生长中光和营养物质利用背后的调节网络。
2 材料方法
太平洋亚历山大藻最初接种于4000细胞/mL放入含有2L经修饰的f/2介质进行处理。三种处理条件分别为高光(HL)、高氮(HN)和对照(CT)。
3 研究思路
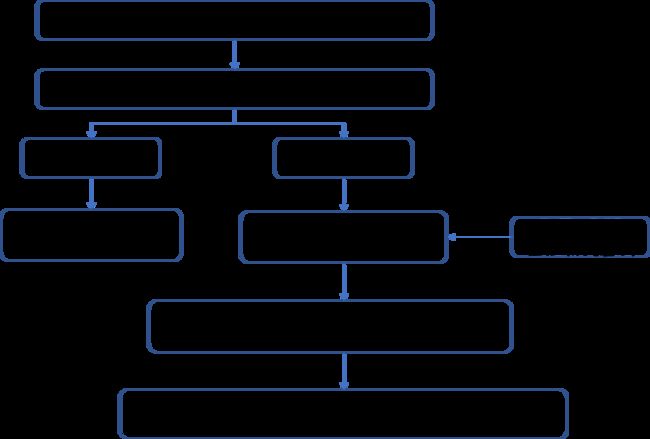
1. 太平洋亚历山大藻在不同条件下的生理反应
为了观察太平洋亚历山大藻的生长状况并确定取样时间,对其在三种处理条件下的生长曲线进行了计数。研究发现太平洋亚历山大藻在第1-3天表现出较低的生长速率,然后在第5-15天快速生长(图1)。15天后,藻类生长趋于稳定。结果表明,高照射和高氮条件对太平洋亚历山大藻的生长有促进作用。选择快速生长期的第12天作为后续实验的采样时间。

图1:不同条件下太平洋亚历山大藻的生长曲线
2. 太平洋亚历山大藻在不同条件下的H3K4me3丰度
在不同处理条件下,太平洋亚历山大藻的H3K4me3的修饰丰度如图2所示。在太平洋亚历山大藻指数快速生长期间,HN和HL条件下H3K4me3的丰度大于CT条件下的丰度。

图2:不同条件下太平洋亚历山大藻H3K4me3丰度的检测
3. 不同条件下太平洋亚历山大藻的H3K4me3相关基因
将获得ChIP-Seq数据与虫黄藻(Fugacium kawagutii)基因组比对,在每种条件下,总共约有2878–7822个peak;发现来自HL的156个基因、来自HN的254个基因和来自CT的803个基因与H3K4me3特异性相关(图3)。在HN和HL条件下,共有36个基因与H3K4me3修饰相关,但在CT条件下没有。
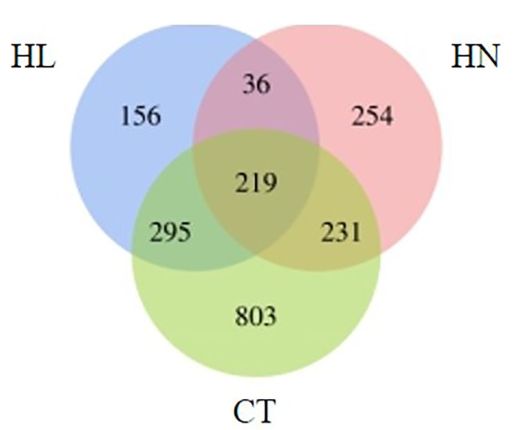
图3:H3K4me3相关基因数量的维恩图
4. ChIP-qPCR
作者又进行ChIP-qPCR以确认ChIP-Seq的结果,如图4所示。ChIP-qPCR共选择了9个参与各种生物过程的基因(来自图3中的156+36+254+219),包括核黄素生物合成蛋白RibBA(RibBA)、真核翻译起始因子2(Euk)、细胞壁α-1,3-葡聚糖合酶mok12(mok12)、核糖体蛋白L14(L14)、泛素激活酶E1(E1)、小亚基核糖体RNA基因(RNA基因),细胞色素b基因(CB)、ATP合酶CF1亚基α(ATP)和18S rRNA基因(18S)。样品免疫球蛋白G用作ChIP-qPCR测定中的阴性对照。与CT相比,大多数选择的基因在HL和HN下表现出显著的富集。每个基因的免疫沉淀DNA和免疫球蛋白G(阴性对照)的相对富集度超过10倍。

图4:ChIP-qPCR在高光照(HL)、高氮(HN)和正常(CT)条件下的结果。
5. 太平洋亚历山大藻HL条件下特有的H3K4me3相关基因的分析
KEGG富集分析表明,与生长和营养吸收相关的途径,如氮代谢、内吞作用、乙醛酸和二羧酸盐代谢、叶酸生物合成、苯丙氨酸代谢和内质网中的蛋白质加工,都显著富集(图5)。在这些富集的途径中,HL条件下特有的H3K4me3相关基因,如硝酸还原酶和谷氨酰胺合成酶,参与氮代谢。此外,一些H3K4me3相关基因,如蛋白26、E3泛素蛋白连接酶和热休克蛋白70(HSP70)参与了内吞途径。

图5:太平洋亚历山大藻HL条件下特有的H3K4me3相关基因的前10个富集KEGG途径
6. 太平洋亚历山大藻HN条件下特有的H3K4me3相关基因的分析
通过KEGG富集分析HN条件特有的H3K4me3相关基因。维生素B6代谢显著丰富。此外,与维生素代谢相关的其他途径,包括叶酸、硫胺素、泛酸和辅酶A生物合成,以及烟酸和烟酰胺代谢,也得到了富集。此外,柠檬酸盐循环、丝裂原活化蛋白激酶信号传导、植物激素信号转导和RNA聚合酶磷酸肌醇代谢途径也得到了丰富(图6)。KEGG分析表明,在HN条件下,太平洋蟾蜍受影响最大的途径是维生素代谢途径。参与维生素B6代谢的磷酸丝氨酸氨基转移酶、外泌体复合物核酸外切酶RRP6和吡哆醇4-脱氢酶三个基因显著富集。

图6:太平洋亚历山大藻HN条件下特有的H3K4me3相关基因的前10个富集KEGG途径
# 结 论 #
太平洋亚历山大藻是一种甲藻,因其快速生长导致大规模开花而臭名昭著。作者旨在从H3K4me3修饰调控的角度,探讨太平洋亚历山大藻在实验室模拟快速生长条件下的分子机制。采用蛋白质印迹法检测在高光(HL)、高氮(HN)和f/2培养基(对照,CT)等不同条件下培养的和平乳杆菌中H3K4me3在快速生长指数期的修饰丰度。结果表明,在HL或HN条件下,H3K4me3的修饰丰度大于CT条件下的修饰丰度。
首次利用染色质免疫沉淀测序技术对不同条件太平洋亚历山大藻进行了H3K4me3的研究。在HL条件下,氮代谢和内吞作用与H3K4me3的调节显著相关。此外,H3K4me3也与HN下的维生素代谢途径显著相关。这些发现表明,H3K4me3在调节太平洋亚历山大藻的快速生长中发挥着潜在的重要作用。这些关于这种甲藻中组蛋白修饰调控网络的知识,为未来相关领域的研究奠定了必要的基础。

图7:高光、高氮条件下太平洋亚历山大藻H3K4me3修饰相关的主要代谢途径