植物DNA甲基化专题 | nature:油棕Karma转座子表观遗传重要发现
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
关于植物DNA甲基化研究,此前已推出见刊于NAR的文章:拟南芥AtHDA6与着丝粒周围DNA甲基化关系研究。 https://mp.weixin.qq.com/s?__biz=MzAwNTY3NDIxMw==&mid=2650648987&idx=1&sn=286fae8141d5e6f1cf26eeab7c8909b2&chksm=83101411b4679d072591d61512e7f9433c3e0c6debda3d93fb8ba6fee487e4f0cf09887a4bf6&token=1026991100&lang=zh_CN#rd
https://mp.weixin.qq.com/s?__biz=MzAwNTY3NDIxMw==&mid=2650648987&idx=1&sn=286fae8141d5e6f1cf26eeab7c8909b2&chksm=83101411b4679d072591d61512e7f9433c3e0c6debda3d93fb8ba6fee487e4f0cf09887a4bf6&token=1026991100&lang=zh_CN#rd
今天要给大家分享关于油棕(oil palm )DNA甲基化的经典研究,本研究成果2015年高分见刊nature。

标题:Loss of Karma transposon methylation underlies the mantled somaclonal variant of oil palm Karma转座子甲基化的缺失是油棕体细胞克隆变体的基础
期刊:Nature
2021影响因子: IF 49.962
发表时间:2015.09.9
方法:EWAS分析、WGBS测序、LDA分析、小RNA
当分化的体细胞重编程为多能性干细胞时,植物和动物中会出现体细胞克隆变异(Somaclonal variation)。在农业领域,体细胞克隆变异阻碍了优良杂交和转基因作物的快速繁殖,但其机制仍然是一个谜。油棕Mantled变异是一种由组织培养引起的体细胞克隆变异,它大大降低了产量,且产油极少,虽然被广泛认为是表观遗传学的影响,但其mantling机制仍然无法解释。
非洲油棕(Elaeis guineensis)是最高产的含油作物,但因为对食用油和生物燃料的需求,以及对热带雨林储量不断减少等可持续性问题的担忧,导致了提高油棕产量的巨大压力。引入tenera杂交品种(dura x pisifera),利用SHELL基因单基因杂交优势将油产量提高了30%。单株高产tenera杂交棕榈(ortets)的克隆株(ramets)提供了更为快速的增产方式,可提高20%~30%。通过未成熟叶片先端组织(“棕榈心脏”)的细胞培养进行快速繁殖,并在补充激素的培养基上进行幼苗再生,可产生数以万计基因相同的克隆棕榈。但与此同时,马来西亚联合种植园的Tan Yap Pau团队注意到克隆株中高频率出现的同源花表型,称为“mantling”,随后Corley研究团队记录了经过长时间培养后出现的有毛棕榈树。在mantled棕榈树中,雌花的退化雄蕊和雄花的雄蕊发育成假心皮(pseudocarpels),通常导致不育的单性结花、果实不育、油产量非常低(图1a-c)。
研究人员使用基于E.guineensis(pisifera)参考基因组DNA芯片进行全基因组DNA甲基化关联分析(EWAS)鉴定与mantled表型相关的表观遗传位点,通过DNA甲基化芯片杂交技术进行对比和统计分析,对每个间隔1-2kb区域测量DNA甲基化密度。全基因组DNA甲基化图谱由单性结实mantled(n=43)、正常ramets(n=54)和ortets(n=10)构建,EWAS分析构建的图谱与棕榈样本全基因组重亚硫酸盐测序(WGBS)分析构建的图谱非常相似(图1d),证明了其可靠性。

图1:EWAS分析
在全基因组分辨率下,ortets和ramets样本DNA甲基化图谱中均一致的在重复序列区域中显示超高甲基化,在数千个基因位点出现差异甲基化(图1d)。其中大部分(~90%)在mantled中DNA低甲基化,与此前报道的5mC含量降低一致。大多数DNA低甲基化基因位点(~75%)是转座子和重复序列,而不太常见的DNA高甲基化基因位点则位于基因序列中。使用从4个独立来源种群获得的15个独立体细胞克隆谱来最大化基因多样性,首先在每个来源种群中鉴定正常和完全mantled样本间的显著差异甲基化区域(DMR),然后对比结果(图1e)。尽管在mantled样本和正常样本之间检测到数万个DMRs,但其中99.9%是4个种群中的一个(94.4%)或2个(5.5%)独有,表明不同种群对组织培养的表观遗传反应存在显著的基因差异。单个芯片检测到所有4个种群中正常和mantled样本之间的差异甲基化(图1e),主要位于EgDEF1区域~35kb内含子5内(图2a),该基因是金鱼草 DEFICIENS(DEF)和拟南芥APETALA3(AP3)基因B类MADS-box转录因子基因的油棕直系同源物。
EgDEF1在12号染色体上跨度约40kb。Ty1/copia反转录转座子(Rider)位于上游,而Ty3/gypsy反转录转座子(Koala)位于内含子5内。与早期研究结果一致,在多个物种的mantled克隆之间未发现这些反转录转座子的DNA甲基化差异。但第三个此前未报道的位于内含子5的重复序列,与水稻Karma LINE元件具有同源性。Karma在水稻胚胎组织培养中被激活,但仅在再生植株中随着该序列的跨代DNA低甲基化持续转座。独特的芯片特性可持续检测到mantled克隆样本中的低甲基化,包括预测的剪接位点(GAACAG^ATGC),在Karma中所有三个额外的芯片比对也检测到mantled克隆样本中的显著低甲基化。(图2a)
为了验证Karma低甲基化,研究人员对包含来自5个独立克隆谱遗传相同的ortets、单性结实mantled、和正常ramets三类样本进行进行全基因组重亚硫酸盐测序(WGBS)。结果显示EgDEF1全基因组中CG甲基化没有变化,但mantled样本中Karma CHG甲基化(H=A,C或T)水平显著降低,揭示了约70个CHG位点的DMR,而CHH甲基化程度要少很多,只是略微降低(图2b)。为验证不相关克隆棕榈中的差异CHG甲基化,研究人员使用定量聚合酶链反应(qPCR)来定量DMR内BbvI和RsaI限制性位点的CHG甲基化(图2c),来自21个克隆谱和4个独立来源的共49个棕榈的panel:8个杂交ortets克隆和13个正常克隆、19个单性结实的mantled克隆、2个可育的mantled克隆和7个回复突变型克隆(revertant clones),分别结出mantled变异和正常的果实。
线性判别分析(linear discriminant analysis,LDA)在mantling检测中具有93%的灵敏度和100%的特异性(图2c),来自所有7个回复突变型棕榈叶被评分为mantled,这与正常束在发育后期出现的观察结果一致。通过跨Karma剪接位点区域的重亚硫酸盐测序进一步分析了两个假阴性mantled棕榈(图2c)。测序结果显示正常克隆株具有密集的CHG甲基化,mantled株CHG甲基化丢失(图2d-e),而假阴性mantled样本在剪接位点附近CHG甲基化丢失(图2f-g),该结果足以预测mantled表型。由于其较强的预测性,研究人员分别将被mantled高甲基化和低甲基化基因分别命名为Good Karma和Bad Karma。
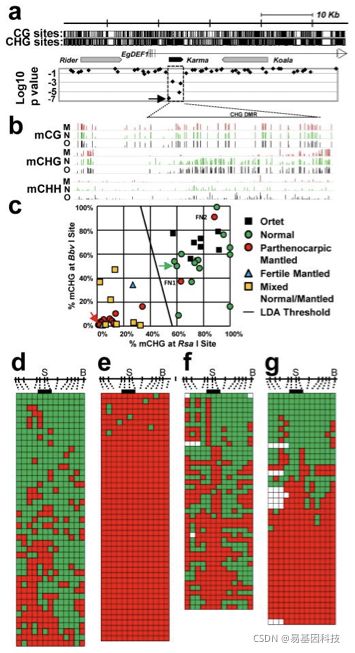
图2:Karma 低甲基化与mantled变异有关
两个谱系的回复突变型棕榈混合束能同时结出正常和mantled变异的果实,类似受转座子调控的玉米的表观基因。第一个谱系包括两个回复突变株,每束分别有99%和95%的正常果实,其中异常果实只有一个或两个小假心皮(图3a-c)。第二个谱系包括只有7%正常果实的mosaic株。在3个分株中,叶片中Bbv I位点的CHG甲基化较低(图3d),与其他回复突变体一致(图2c)。两个回复突变株的果实中DNA甲基化状态恢复,而mantled变异mosaic株中甲基化状态没有恢复(图3d-f)。与玉米、Linnaria和其他植物研究结果一致,强有力的证实了Karma低甲基化是mantled变异的原因所在。然而本研究未观察到单个mantled果实和正常果实之间的差异甲基化,这可能反映了B类同源的非细胞自主性(图3a-d),同样该结果也在金鱼草和拟南芥中观察到。对正常和mantled果实的重亚硫酸盐测序分析结果显示出剪接位点的高甲基化和低甲基化reads(图3e-f),表明这些果实确实是高甲基化和低甲基化细胞的嵌合体。

图3:回复突变型棕榈中的KAMA甲基化
剪接位点附近的DNA甲基化会影响可变剪接,为评估可变剪接,研究人员基于转录组测序(数据未显示)建立EgDEF1转录本并通过RT-PCR验证。如此前报道,在正常和mantled的花中发现两种形式的EgDEF1转录本:全长EgDEF1转录本(cDEF1)和提前终止的转录本,包括外显子1-5和221bp的内含子5(tDEF1),此外在mantled雌花中鉴定了第三种替代转录本kDEF1。(图4a)

图4:可变剪接和24nt小RNA的丢失
另外,研究人员在茎尖分生组织(第0阶段)和未成熟雌花发育的4个阶段(第1-5阶段)中评估了三个转录本的表达(图4b)。结果显示kDEF1表达明显受限于mantled(但不正常)雌花的第3-5阶段,强烈暗示在mantled中的转录调控作用。相比之下,在mantled雌花中,cDEF1仅检测到(尽管显著)较低水平的表达,而tDEF1没有变化(图4b)。此前报道的每个阶段中tDEF1和cDEF1转录差异可能是kDEF1转录的结果。
在植物中,24nt小干扰RNA(small interfering RNA,siRNA)指导non-CG甲基化,研究人员在茎尖分生组织(第0阶段)中鉴定出了反义链24nt Karma siRNA簇,它们在mantled中降低或丢失(图4c)。在正常和异常克隆棕榈的多胚胎组织培养中,小RNA(small RNA)经历了从24nt至21nt的动态变化。Karma甲基化和24nt siRNA(图4c)在正常培养中可减少2-7次传代,而在异常培养中丢失,表明了mantled变异的起源:如果分生组织是24nt siRNA的来源(图4c),从分生组织分离的叶细胞在培养中随时间的推移而出现24nt siRNA丢失和non-CG DNA甲基化。
mantling在过去的30年中一直是分子遗传学研究难以捉摸的研究对象。研究已经证实mantled性状是B类MADS-box EgDEF1基因内Karma转座子表观遗传修饰的结果,并将其命名为MANTLED。kDEF1转录本在变化过程中的出现(图4b)表明,Karma转座子甲基化的缺失是油棕体细胞克隆变体的基础。
易基因小结
本研究使用表观全基因组关联分析(Epigenome Wide Association Studies, EWAS)来鉴定Mantled变异的候选基因。与水稻Karma相关的LINE反转录转座子在同源基因DEFICIENS内含子区域的DNA低甲基化,在所有Mantled克隆基因中均出现,且与可变剪接和提前终止相关。Karma剪接位点周围区域密集的甲基化(Good Karma基因)则预示正常坐果,而低甲基化(Bad Karma基因)则预示同源转化(homeotic transformation)、单性结实(parthenocarpy)和产量骤降。组织培养中Karma甲基化和小RNA的丢失证实了mantled变异起源,这种mantled变异并不遵循孟德尔遗传规律。本研究结果揭示了在油棕幼苗阶段可以预测和剔除mantled变异,有助于提高产量、优化环境敏感的土地资源。
文献来源:10.1038/nature15365
全文解读:植物DNA甲基化专题 | nature:油棕Karma转座子表观遗传重要发现https://mp.weixin.qq.com/s?__biz=MzAwNTY3NDIxMw==&mid=2650649174&idx=1&sn=de2c2163f0803b39eaac72b26a82326e&chksm=831017dcb4679eca02867044d795642e29f96138f2199802d7edac0e32db7ab14f4c9e0ae532&token=1026991100&lang=zh_CN#rd