(1)背景知识
真核生物RNA可以携带100多种化学修饰,其中RNA甲基化修饰约占60%,而N6-甲基腺嘌呤(m6A)在甲基化修饰中最为普遍,占有率高达80%。从2017年至今,RNA m6A研究热度不减,已有7篇在《Nature》、2篇在《Cell》、2篇在《Cancer cell》上发表。
概念
m6A是腺苷酸上第六位N上发生甲基化,主要存在于mRNA的CDS区与3’ UTR区,影响mRNA的稳定性、翻译效率、可变剪切和定位等。此外,LncRNA、microRNA、circle RNA等非编码RNA也存在m6A位点。
分子机制
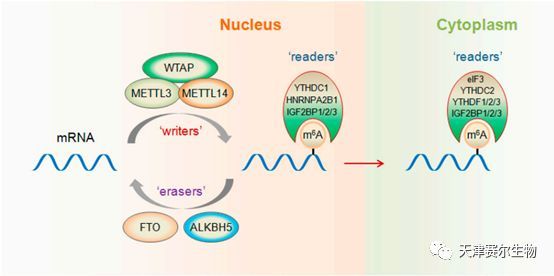
mRNA RRACH序列中的腺嘌呤在甲基化酶的作用下,其第6位N上的H被CH3取代发生甲基化,此过程是可逆的;甲基化的mRNA与m6A识别蛋白结合,进而影响细胞核内miRNA初级转录本的加工、miRNA前体的剪切、mRNA的运输及细胞质中mRNA的翻译或降解。
核心分子
writers:甲基化酶,介导RNA甲基化修饰过程,目前已知的成分有METTL3、METTL14、WTAP和KIAA1429
erasers: 去甲基化酶,有FTO和ALKBH5
readers: m6A结合蛋白,识别并结合mRNA上的m6A位点,有YTH结构域蛋白(YTHDC1、YTHDC2、YTHDF1、YTHDF2、YTHDF3)、核不均一蛋白(HNRNPA2B1)和胰岛素样生长因子(IGF2BP1、IGF2BP2、IGF2BP3)
(2)文献
Gene signatures and prognostic values of m6A regulators in clear cell renal cell carcinoma - a retrospective study using TCGA database. (Aging; 生物二区;IF:5.179;)
摘要
m6A是mRNA修饰的最常见形式。然而,关于其在透明细胞肾中的作用知之甚少。本研究旨在确定m6A调节因子的基因特征和预后价值肾透明细胞癌。在这项研究中,使用来自TCGA数据库的总共528名ccRCC患者的CNV数据。使用对数秩检验和Cox回归模型进行生存分析。使用卡方检验m6A调节基因的改变与临床病理学特征之间的关系。结果显示,m6A调节因子的改变与病理分期有关。m6A调控基因的任何CNV改变都比具有二倍体基因的具有更差的OS和DFS。而且,m6A“writer”基因缺失是OS的独立危险因素,“eraser”基因的拷贝数增加可能以协同方式放大“writer”基因缺失的效果。此外,‘writer’基因METTL3的低表达与脂肪生成和mTOR通路。因此,我们首次确定了遗传改变m6A调节因子在ccRCC中与临床恶化之间的显着关系特点。这些发现为我们了解ccRCC中RNA的表观遗传修饰提供了线索。
介绍
肾细胞癌(RCC)是最致命的泌尿生殖系统肿瘤。最新的癌症统计报告表明每年在美国有超过65,000例新病例,造成近15,000人死亡,这使它成为男性中第六最常见的肿瘤。透明细胞癌(ccRCC)是细胞癌最常见的类型(80%)。临床上,高达16%的ccRCC患者诊断时已发生转移,并且5年存活率仅为12%。目前肿瘤学和外科学的发展只是略有改善ccRCC的预后。至于局限性的ccRCC,20%至30%的患者治疗后会出现复发,而且目前尚没有已经被批准的治疗方案用于降低复发,进展或死亡。近年来,虽然靶向治疗被证明可以延长转移患者的生存期,但是患者中位生存期仍不到3年。此外,耐药性和经济负担是临床实践中面临的相当大问题。因此,探索ccRCC的发病机制和新的治疗靶点是研究热点。
DNA和DNA表观遗传改变组蛋白已在肿瘤进展中得到广泛研究并导致了许多治疗方法的发展,包括组蛋白去乙酰化酶抑制剂和靶向缺氧相关通路的药物。距离从上述两个分子,RNAs的作用不同的细胞过程引起了广泛的关注,并在过去十年成为一个快速发展的领域。超过一百种核苷酸存在于不同类型的RNA中,包括rRNA,tRNA,mRNA,snRNA等。修饰的RNA,尤其是mRNA,具有重要意义在基因的转录后调控中的作用表达。在真核生物中,m6A是最常见的mRNA修饰的形式,丰富的已发现腺苷总量为0.1-0.4%残留物。 m6A被证明在整个转录组普遍存在,实际上在超过7,600个基因的mRNA和超过300个非编码RNA 存在m6A。一般来说,m6A在人与小鼠高度保守,位于3'UTR,围绕终止密码子和内部外显子,其功能是改变RNA稳定性,剪接,细胞内分布和翻译。该细胞m6A状态由一组基因介导被称为“writer”(WTAP,METTL3和METTL14),“eraser”(FTO和ALKBH5)和“reader”(YTHDF1,YTHDF2,YTHDF3,YTHDC1和YTHDC2)。“writer”形成一个甲基转移酶复合物和上调m6A水平,而“eraser”是m6A脱甲基酶,与“writer”功能相反。此外,“reader”是解码的效应器m6A甲基化信息并将其转化为功能信号。
m6A失调涉及多种细胞过程并导致细胞增殖减少,自我更新能力受损,发育缺陷和细胞死亡。据报道,m6A调控基因的改变起着重要作用在各种人类疾病的发病机制中的作用包括肥胖,精子发生受损,神经元疾病和免疫疾病。更多最近,m6A调控基因的改变了已被证明可以促进两个乳房的进展通过癌症治疗癌症和血液系统恶性肿瘤干细胞形成和异常分化状态维护。 另一项研究也证明了这一点METTL3,一种主要的RNA N6-腺苷甲基转移酶,促进肝癌进展通过YTHDF2依赖的转录后沉默SOCS2 。 虽然发现了m6A与不同类型的肿瘤发生有关癌症,关于它们之间的关系知之甚少m6A相关基因和ccRCC。 因此,在这项研究中,我们分析ccRCC的临床和测序数据来自TCGA的队列,并评估改变ccRCC中10个m6A调控基因的谱图作为遗传改变与遗传改变之间的关联临床病理特征包括生存。
结果
-
m6A调控基因的突变和CNV
在451例sccRCC患者测序数据中,m6A调控基因突变仅在19个独立样本(Table1); 然而在具有CNV数据的528个ccRCC样品中,观察到m6A调节基因具有高频的CNV(Fig. 1A)。 例如“reader”基因YTHDC2最高其CNV事件的频率为(55.11%,291/528),其次METTL3(30.11%,159/528),它们是am6A调控基因中的“reader”和“writer”基因。 此外,我们也观察到频繁的VHV(89.02%)和TP53的CNV(14.58%),该结果与已发表的文献一致。

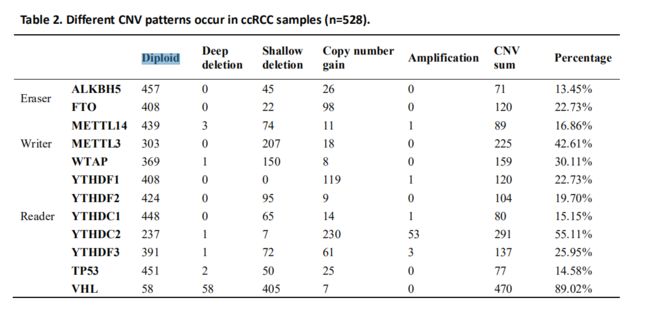
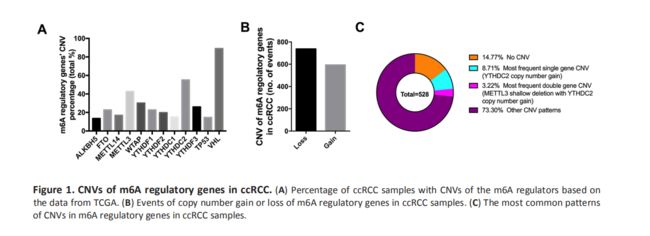
接下来,我们评估了ccRCC样本中的CNV模式并发现大多数CNV事件的结局是拷贝数缺失(737/1331)(Figure 1B, Table 2),这与AML中的CNV状态相似。 YTHDC2基因的拷贝数数量增加是最常见的m6A调节基因C拷贝数改变(Figure 1C),同时METTL3和YTHDC2也在双基因CNV中排名第一,暗示着m6A编码基因在RNA过程中的重要性m6A修改。
-
m6A调控基因的改变与临床病理学和分子特征有关。
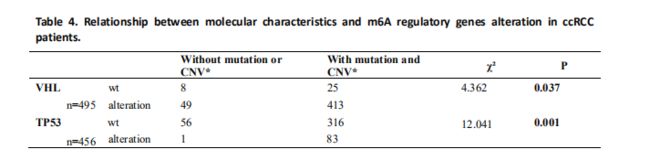
我们评估了m6A调控基因的改变(CNV和/或突变)与患者的临床病理学特征之间的关系。 结果显示,m6A调节基因的改变与更高的‘Fuhrman Nuclear Grade’显着相关(Table 3)。 由于VHL和TP53在发病机制中起重要作用对于ccRCC ,我们进一步评估了m6A调控基因的变异是否与这两个基因的改变有关。 正如所料,m6A调控基因的改变与VHL和TP53的改变显着相关; 事实上,在57例TP53改变患者中,m6A调节基因的改变中只有1个样本缺失(Table 4)。接下来评估了m6A调节基因改变对mRNA表达的影响。 结果显示,在525个ccRCC样品中,mRNA表达水平与不同的CNV模式显着相关。 对于所有10个调节基因,拷贝数增加与较高的mRNA表达有关; 而深缺失或浅缺失则导致mRNA表达下降(Figure 2)。 此外,我们对含有约130对ccRCC组织和正常组织的组织中的METTL3和METTL14蛋白进行了一些免疫组织化学染色,以证实这些发现。 结果与我们的分析一致,METTL3和METTL14在ccRCC组织中高于正常表达(P <0.05)(Fig S1)。 然而,由于缺乏预后数据,我们无法在本研究中进行生存分析。

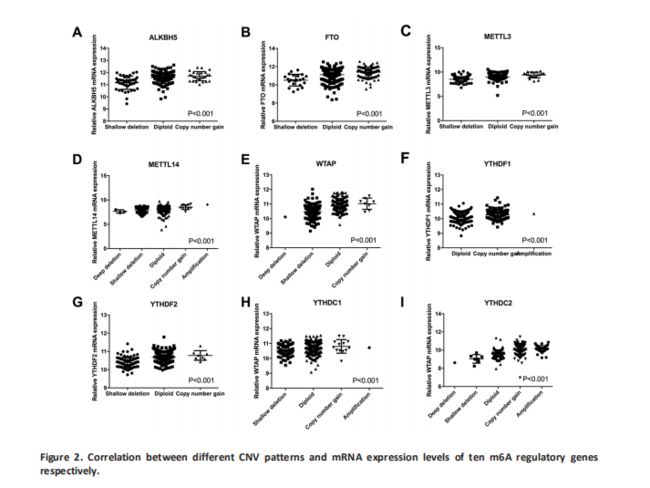
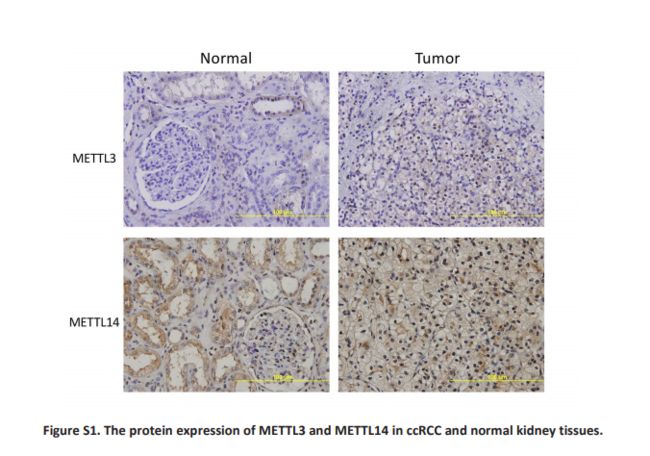
-
m6A调节基因的CNV与ccRCC患者的存活之间的关联
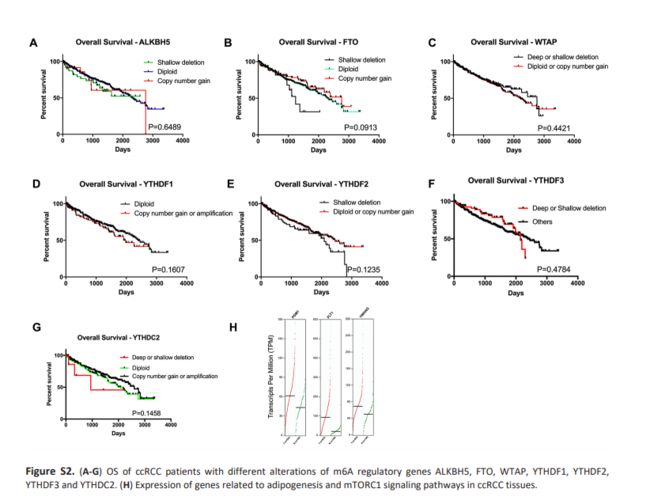
为了探讨CNVs在m6A调节基因中的预后价值,我们分析了CNVs对ccRCC患者的总生存期(OS)和无病生存期(DFS)的影响,并发现发生m6A调节基因CNVs的个体和没有发生m6A调节基因CNVs的个体,在OS和DFS没有差异(Fig 3A-B)。此外,对10个基因的单独分析显示受YTHDC1,METTL14或METTL3缺失影响的患者(m6A的一个‘reader’和两个‘writer’基因)具有较差的OS和DFS(Fig 3C-H); 而基于其他10个m6A调节基因的CNV,在不同亚组之间未观察到显着差异(Fig S2)。多变量Cox回归分析表明,m6A调节基因的改变是总体存活的独立危险因素(Table5)。考虑到‘writer’基因是一组甲基转移酶并且是m6A调节过程中最重要的部分,结果表明m6A水平的下调可能与患者存活率差有关。
为了证实上述结论,我们接下来在两种CNV影响的患者中进行了测试(‘writer’基因的缺失和‘eraser’基因的拷贝数增加)。结果显示,与仅具有‘writer’基因缺失的那些相比,同时具有‘eraser’基因的拷贝数增加的‘writer’基因缺失的患者具有更差的OS和DFS(Fig 4A-B)。这为下调的m6A水平与较差的预后之间的关系提供了更多证据。
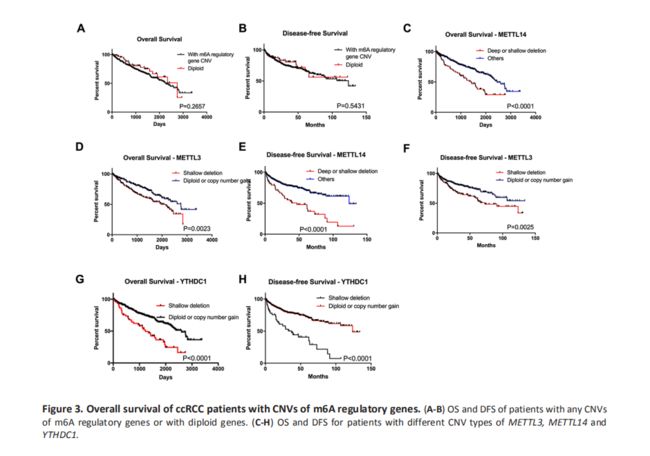

-
METTL3拷贝数丢失的功能富集分析
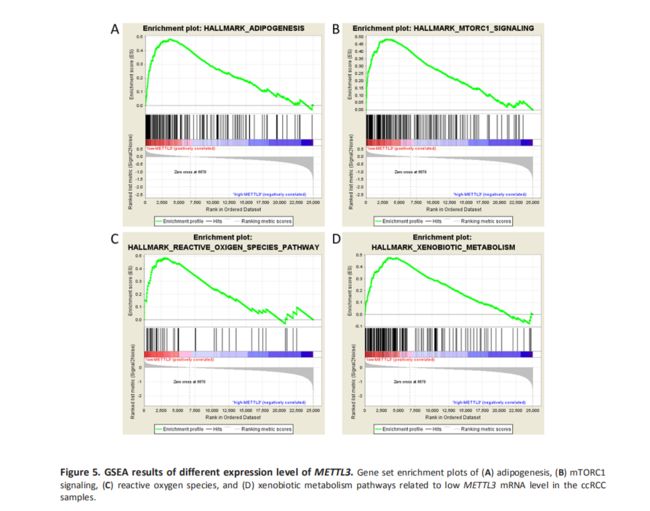
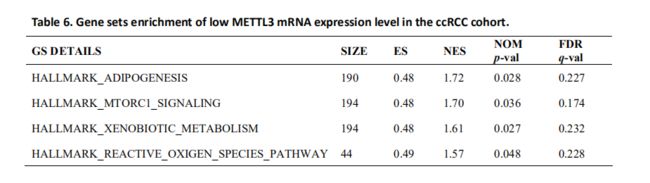
鉴于METTL3在甲基化过程中的重要性和我们的结果,我们决定探讨m6a失调在ccRCC发病机制中的作用。我们检验了不同METTL3 mRNA表达水平样品中的富集基因集。基因富集分析表明低的METTL3表达与脂肪形成、mTOR通路和活性氧(ROS)等重要的生物学过程有关(Table6和Fig 5),提示了ccRCC发病的潜在机制。为了验证我们的发现,我们检查了与上述途径相关的基因表达。我们发现在RCC肿瘤组织中有三个与脂肪形成和mtorc1信号通路的基因高表达(Fig S2-H),该结果证实了GSEA的结果。此外,一些研究发现METTL3可以参与脂肪形成和mTORC1信号传递途径,这与我们的结果是一致的。需要进一步的研究来说明METTL3和METTL4对下游基因的调控作用

材料和方法
-
伦理声明
所有临床数据,CNV,突变,mRNA表达数据均由cBioportal平台和TCGA-assembler 从TCGA网站中检索下载到,这些数据向公众开放。因此,确认所有书面知情同意书均已达成。
-
数据处理
在TCGA数据库中,我们确定了528名患有CNV数据和病理报告的ccRCC患者。对于CNV,使用分段分析和GISTIC算法识别拷贝数变化的loss和gain水平。为了研究CNV和/或突变状态的临床病理学意义,将该ccRCC队列分为两个亚组;:“具有这10个M6A调节基因的突变和/或CNV”和“没有CNV和突变”。从RNA-Seq V2 RSEM计算mRNA表达数据,并在分析mRNA表达与CNV之间的关系之前应用Log scale。
-
基因集富集分析(GSEA)
GSEA由JAVA程序与MSigDB v6.1提供,并从网站下载Broad Institute 。在该研究中,根据METTL3表达水平的第一和第四四分位数将病例分成两组。共计18419个基因参加了GSEA分析。本研究中使用了Hallmark基因集“.all.v6.0.symbols.gmt”。统计学标准:标准化p值<0.05,并且错误发现率(FDR)<0.25被认为显着富集。
-
统计分析
使用SPSS 20.0(IBM,Chicago,USA)和GraphPad Prism 6.0(GraphPad Software,La Jolla,CA,USA)分析所有统计数据和图。用卡方检验或Mann-Whitney U检验分析m6A调节基因-CNV与临床病理学特征之间的关联。 Kaplan-Meier曲线和对数秩检验用于评估m6A调节基因的改变的预后值。使用SPSS进行Cox比例风险回归模型。 p值<0.05的所有统计结果被认为是显着的