Single-Cell Sequencing of Peripheral Mononuclear Cells Reveals Distinct Immune Response Landscapes of COVID-19 and Influenza Patients

https://www.sciencedirect.com/science/article/pii/S1074761320303162#mmc1
影响因子: 22.553PMID:32783921
期刊年卷:Immunity 2020 09 15;533(3)
医学一区 免疫学 Q1 4/155
DOI:10.1016/j.immuni.2020.07.009
亮点:
作者在 COVID-19 和流感患者中生成了 PBMC 单细胞图集
COVID-19 和流感患者的血浆细胞显著增加
COVID-19 具有 XAF1,TNF-α和FAS诱导的T细胞凋亡的特征
与流感病毒(STAT3 /NFκB)相比,COVID-19激活了独特的途径(STAT1 / IRF3)
2019年冠状病毒病(COVID-19)大流行构成了当前全球范围内的公共卫生威胁。但是,与其他传染病相比,人们对其标志的了解还很少。在这里,作者报告了在COVID-19和甲型流感病毒(IAV)感染的患者中,纵向收集的外周血单个核细胞(PBMC)的单细胞转录态势。作者观察到COVID-19和IAV患者的血浆细胞增加,而XIAP相关因子1(XAF1),肿瘤坏死因子(TNF)和FAS诱导的COVID-19患者T细胞凋亡增加。进一步的分析显示,COVID-19(STAT1和IRF3)与IAV(STAT3和NFκB)患者相比,激活的信号通路不同,并且关键因子的表达存在明显差异。与 IAV 患者不同,IL6R 和 IL6ST 的表达在 COVID-19 患者中被上调,这在发病过程中协同促进促炎细胞因子的增加。作者还发现,几个干扰素(IFN)刺激的基因(ISG;包括 ISG15,IFI44,IFI44L 和 RSAD2)在 COVID-19 患者的 PBMC 中被特异性上调,增强了病毒感染后的抗病毒和免疫调节功能。支持了COVID-19患者中促炎细胞因子增加的临床观察。因此,作者提供了PBMC的概况,并揭示了COVID-19和IAV患者的独特免疫应答途径。
结果
COVID-19 和 IAV 患者的 PBMC 的单细胞转录景观
为了调查 COVID-19 患者中 SARS-CoV-2 感染的发病机理和机制,从三个健康对照,两名 IAV 感染患者和五名 COVID-19 患者(包括四名患者(COV-1 至 COV) -4)病程简单,一名患者(COV-5)随后病情加重(图1A;表S1)。选择季节性感染 IAV 的患者作为对照,因为 SARS-CoV-2 和 IAV 均为传染性 RNA 病毒,均引起呼吸道感染。但是,与 IAV 感染相比,COVID-19 具有明显的临床特征,包括高发病率和高死亡率。根据阳性核酸检测结果,在症状发作的 5-10 天内招募了 COVID-19 患者,并将首次收集 PBMC 的日期命名为第 1 天。表S1中列出了与疾病发作和样本收集相对应的时间点。如图1A 所示,在随后的时间点也收集了 PBMC。过滤出低质量的细胞后,作者从 46,022 个细胞中获得了转录组数据集,每个时间点每个参与者平均有 2,000 个细胞(图1B)。为了发现 COVID-19 中的免疫细胞群,作者进行了无监督的聚类并获得了 15 个细胞群(图1C)。根据经典细胞类型标记物的表达,可以鉴定出免疫细胞,例如 T 细胞,B 细胞,单核细胞,自然杀伤(NK)细胞,DC,干细胞和巨核细胞(图1C)。通过这种方法,5 个群体注释为 T 细胞,包括幼稚 T 细胞(CD3+CCR7+GZMA-),活化的 CD4+T 细胞(CD3+CD4+IL7R+),细胞毒性 CD8+T 细胞(CD3+CD8+GZMA+),粘膜相关不变性 T 细胞(MAIT 细胞;CD3+SLC4A10+)和循环性 T 细胞(CD3+MKI67+);四个群体注释为 B 细胞,包括幼稚 B 细胞(MS4A1+IGHG1-),浆细胞(MZB1+IGHG1+),循环血浆细胞(MZB1+IGHG1+MKI67+),和记忆 B 细胞(MS4A1+IGHG1+) ; 其中两个被标注为 NK 细胞(KLRF1+);一个种群被标记为 DC(CD1C+LYZ+)和单核细胞(LYZ+CD68+)(图1D 和S1A)。大多数簇由多名患者的细胞组成,表明患者之间具有共同的免疫特征。此外,来自患者的 PBMC 样本不表达ACE2和TMPRSS2受体,并且不显示病毒读数,表明 SARS-CoV-2 可能不会感染 PBMC(图S1B)。
下载:下载高分辨率图片(1MB)下载:下载全图
图 1 参与者的 PBMC 衍生的免疫细胞的单细胞基因表达谱
(A)研究设计的示意图。这项研究包括 10 名受试者,包括 3 名健康捐献者,5 名 COVID-19 患者和 2 名受 IAV 感染的患者。
(B)条形图显示了在不同时间点每个供体的每个样品的 log10转化细胞数。蓝色代表三位健康的供体,橙色代表两名被 IAV 感染的患者,五名使用五种不同颜色显示的 COVID-19 患者。
(C)来自十个供体的 46022 个细胞的聚类结果。每个点代表一个单个细胞,根据细胞类型进行着色。巨核细胞。
(D)细胞类型簇中细胞分型基因的表达水平。CD3G 指示 T 细胞,KLRF1 和 XCL1 指示 NKs,MS4A1 指示 B 细胞,IGHG1 和 MZB1 指示浆细胞,CD68 指示单核细胞,LYZ 指示 DCs,MKI67 和 TOP2A 指示循环的 T 细胞,GZMA 指示细胞毒性 CD8+T 细胞和 NKs,和 PPBP 表示巨核细胞。
另请参见图S1和表S1。

COVID-19 和 IAV 患者的 PBMC 中的浆细胞增加
PBMC 人群的一般模式在患者之间是可比的(图2A;表S2)。在感染 COVID-19 和 IAV 的患者中,浆细胞和循环浆细胞的比例均显著增加(图2B 和S2),这两种类型的病毒感染之间没有差异。增加的浆细胞可以诱导保护性中和抗体,以防止病毒感染细胞。不出所料,当检查 COVID-19 患者 B 细胞中上调基因的功能与健康供体相比时,蛋白质复合物装配和蛋白质运输相关途径尤其丰富,这可能是因为在此期间合成了大量蛋白质处理(图2C)。B 细胞激活相关的途径也得到了丰富(图2C),代表性基因包括PRDM1,XBP1和IRF4(图2D)。浆细胞的身份和功能取决于转录因子PRDM1,XBP1和IRF4(Ochiai等人,2013;Shaffer等人,2004)。PRDM1在确定和塑造成熟 B 细胞分化的分泌臂并促进免疫球蛋白(Ig)合成中起着核心作用。XBP1在 CREB-ATF 家族中,它是一种正向转录因子,在浆细胞中大量表达,对于增加浆细胞中的蛋白质合成至关重要(Shaffer等,2004)。IRF4调节免疫球蛋白类别转换的重组,并且持续且较高浓度的IRF4可以促进浆细胞的生成(Ochiai等人,2013)。继浆细胞增加和相关转录因子的表达后,作者发现与健康对照组相比,活化的 CD4+T 细胞上CD2AP的表达升高(图2E)。CD4+中的衔接子分子 CD2APT 细胞调节滤泡辅助性 T 细胞的分化并改善病毒感染中的保护性抗体反应(Raju等人,2018)。除了支持浆细胞功能外,在活化的 CD4+T 细胞和细胞毒性 CD8+T 细胞中,TNFSF14 的表达也增加了。这些因子促进 T 细胞活化以及 T 细胞从外周血中募集到组织中。KDM5A编码 NK 细胞和 T 细胞活化所需的 H3K4me3 脱甲基酶,在 NK 细胞和 COVID-19 的细胞毒性 CD8+T 细胞中被上调(图2E)。综上所述,这些结果表明,在 COVID-19 患者中浆细胞的升高以及 T 细胞和 NK 细胞活化的增强可能有助于抵抗病毒。
下载:下载高分辨率图片(1MB)下载:下载全图
图 2. SARS-CoV-2 感染期间免疫细胞的动态组成和功能变化
(A)每个样品中的细胞类型频率。条形按细胞类型着色。
(B)来自健康供体(Ctrl)(n = 3),COVID-19 患者(COV)(n = 16)和 IAV 患者(IAV)(n = 4)的样本之间血浆和循环血浆比例的差异。使用学生 t 检验来检验差异的显著性。p <0.05,p <0.01,**p <0.001。
(C)与 B 细胞中健康对照相比,COVID-19 患者中上调基因的 GO 术语富集。
(D)健康供体(Ctrl)和 COVID-19 患者(COV)在浆细胞中 B 细胞激活相关基因PRDM1,XBP1和IRF4的差异表达水平。
(E)健康供体和 COVID-19 患者样品中活化的 CD4+T 细胞,细胞毒性 CD8+T 细胞和 NK 中 T 细胞活化相关基因的表达水平。在上图中,每个点的颜色代表基因的表达水平,而点的大小代表在特定细胞类型中表达基因的细胞比例。在下部面板中,使用学生 t 检验测试了健康捐献者(Ctrl)(n = 3)和 COVID-19 患者(COV)(n = 16)之间的差异。p <0.05,p <0.01,**p <0.001。
另请参见图S2和表S2。

COVID-19 患者的 IFN 反应和淋巴细胞凋亡
进行基因本体论(GO)分析以深入了解 COVID-19 患者和健康对照之间不同细胞亚群的功能。“对 I 型 IFN 信号的响应”组中的基因富含 D1 和 D4 的 T,B 和 NK 细胞亚群,但不包含 D16 样品(图3A 和S3A),这与 IFN 反应是对于病毒感染引发的反应至关重要。一致地,“对病毒信号的防御反应”基因也在 D1 和 D4 而非 D16 的所有五名 COVID-19 患者的 T,B 和 NK 细胞中富集(图3A 和S3)A),表明持续不断的针对 SARS-CoV-2 病毒的免疫反应。此外,内质和蛋白质解折叠相关的途径在三个时间点(D1,D4 和 D16)都特别富集于 B 细胞中(图3A 和S3A),这可能是由于血浆比例更高由于抗体生产过程中对蛋白质合成的需求很高,因此 B 细胞簇中的 B 细胞具有很高的免疫力。SARS-CoV-2 感染中的其他信号传导途径,例如“染色体组织的调控”,“ DNA 构象变化”等也被上调。这些基因的作用需要进一步研究。
下载:下载高分辨率图片(2MB)下载:下载全图
图 3 COVID-19 患者与 IFN 应答和细胞凋亡相关基因的分析
(A)与不同细胞群体中的健康对照相比,COVID-19 患者第 1 天样品中通过 GO 分析获得的前 20 个丰富的生物学过程。点颜色表示富集的统计显著性(p),点大小表示每个术语均注明的基因比例。
(B)与不同细胞亚群中健康对照相比,来自 COVID-19 患者的第 1 天样品中差异表达的基因。红点代表 COVID-19 患者的上调基因(调整后的 p <0.01 且倍数变化(FC)≥2),而蓝点代表COVID-19患者的下调基因(调整后的p <0.01和FC≤0.5)。log2(FC)≥1.5的基因用基因符号标记。
(C)ISG15,IFI44L,MX1和XAF1在健康供体(Ctrl)(n = 3)和 COVID-19 患者(COV)(n = 16)中的基因表达。使用学生 t 检验来检验差异的显著性。p <0.05,p <0.01,**p <0.001。
(D)基因通过 mfuzz R 包装沿着疾病的进展按其表达模式聚集。
(E)GO 分析显示,每个基因簇中的前 10 个富集的生物过程。
(F)T 细胞中 COVID-19 患者(COV)(n = 5)和健康对照(Ctrl)(n = 3)之间凋亡相关基因表达水平的差异。应用了学生 t 检验。p <0.05,p <0.01,**p <0.001。
另请参见图S3和表S3。
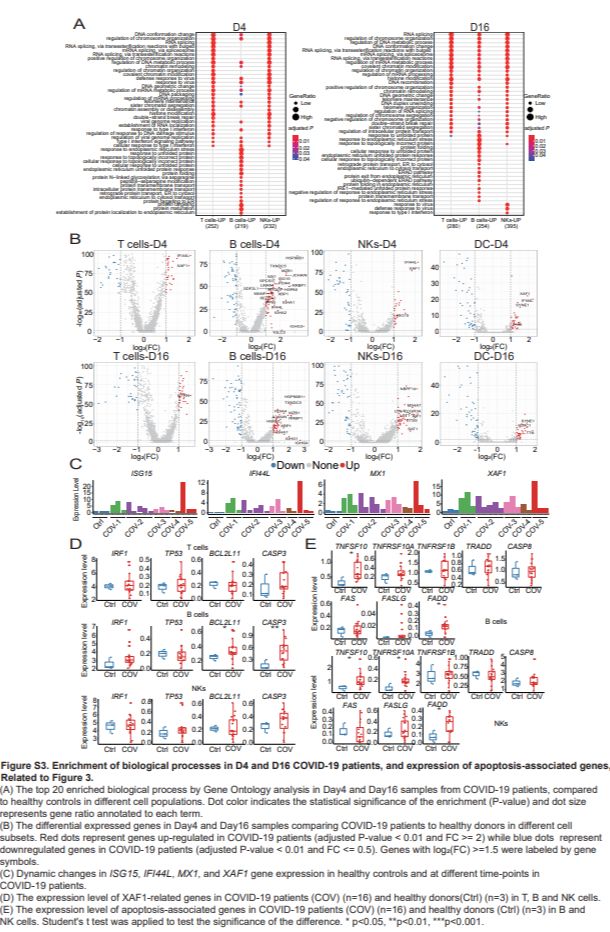
然后在 COVID-19 患者和健康对照之间比较这些转录组谱中的差异表达基因(DEG)。在 D1 和 D4 感染 SARS-CoV-2 的患者中鉴定出了对早期病毒控制至关重要的 ISG(Schoggins和Rice,2011年)(图3B 和S3B),这与“response to type I IFN signaling"/GO分析中的“ I型IFN信号转导”途径相符(图3A 和S3A)。在这些 ISG 中,ISG15,IFI44L,MX1和X 连锁的凋亡抑制因子(XIAP)相关因子 1(XAF1)在 T,B,NK 和 DC 细胞亚群中上调(图3B 和S3B)。与健康对照相比,这四个基因在 COVID-19 患者中的表达水平也要高得多(图3C)。然后,作者在疾病过程中检查了这些基因的转录动力学。为此,作者将 COVID-19 患者从症状发作到出院的疾病过程分为四个阶段(表S3)。作者鉴定了六个时间依赖性表达模式(图3D)并研究了它们的生物学意义(图3E)。聚类 3 包含 158 个基因,这些基因随着时间的推移表达水平下降。这些基因的功能在与 IFN 反应相关的生物学过程中显著丰富,表明这些基因的转录调控是动态的,并且它们在早期时间点被激活而在晚期时间点被去激活(图3E)。簇 1 包含 38 个基因,其表达水平从第 2 阶段开始升高。GO 富集分析表明其功能在翻译和蛋白质合成相关途径中显著富集。这与抗体生产的时机一致(Thevarajan等,2020),因为在此过程中会发生大量蛋白质合成(图3E)。
重度患者(COV-5)对 SARS-CoV-2 感染后对 IFNa 的反应比轻度患者强。此外,ISG15,IFI44L,MX1和XAF1 的表达在疾病进展的较早时间点较高,然后在以后的各个阶段分别降低(图S3C),说明它们对干扰素的动态反应。
泛素样蛋白 ISG15,IFI44L 和 MX1 均在抗病毒反应中起作用(Perng和Lenschow,2018年),而 XAF1 参与促凋亡反应,并且据报道与 IRF-1 形成正反馈回路,从而驱动压力下的细胞凋亡(Jeong等,2018)。XAF1 还通过翻译后修饰增强了 TP53 介导的凋亡(Zou等,2012),因此,分析了与 XAF1 介导的凋亡相关的基因,包括IRF1,TP53,BCL2L11和CASP3(图S3)D)。IRF1,TP53和CASP3 的表达与对照组相比,COVID-19 患者的 T,B 和 NK 细胞亚群的 Bcl2 升高,而BCL2L11在不同细胞亚群中表现出不同的模式。除了 XAF1 相关的凋亡途径外,还在 COVID-19 患者和健康对照者中检查了其他与凋亡相关的途径中的基因表达,包括 TNF 和 Fas 途径(Elmore,2007年)(图3F 和S3E)。相对于健康对照,COVID-19 患者的 T 细胞中TNFSF10(TRAIL)及其受体TNFRSF10A的表达增加。其他 TNF 通路成员,包括TNFRSF1B,在 COVID-19 患者中也相对上调。至于 FAS 途径,在 COVID-19 患者的 T 细胞中,FAS,FASLG和FADD的表达上调,尽管不显著(图3F)。在 B 和 NK 细胞亚群中,在 COVID-19 患者中TNFSF10和FADD显著增加,其他基因轻度升高,除了 B 细胞中的Fas和 NK 细胞中的TRADD显著降低(图S3E)。综上所述,作者发现与 XAF1,TNF 和 Fas 途径相关的上调基因可能导致 COVID-19 患者的 T 细胞凋亡增加。
与 IAV 患者相比,COVID-19 患者的免疫分子特征
接下来,作者试图鉴定与 COVID-19 和 IAV 感染相关的免疫分子标记。因此,作者比较了 COVID-19 患者,IAV 患者和健康对照者的 T 细胞亚群,NK 细胞和 DC 中细胞因子,细胞因子受体和转录因子的表达(图4A 和S4A)。正如基因聚类模式所暗示的那样,COVID-19 患者中上调的基因大多编码促炎性细胞因子,细胞因子受体和 IFN 反应性转录因子,而在 IAV 患者中,促炎性转录因子和与病毒相互作用的宿主因子似乎高度表达。
下载:下载高分辨率图片(1MB)下载:下载全图
图 4 与 IAV 相比,COVID-19 的标志(通过细胞因子,细胞因子受体和转录因子的单细胞分析显示)
(A)在活化的 CD4+T 细胞群体中,COVID-19 患者,IAV 患者和健康对照中关键细胞因子,细胞因子受体和转录因子的相对表达水平(Z评分)。
(B)在 IAV 患者的活化 CD4+T 细胞中高度表达的四个代表性基因的表达水平。上图:点图中每个点的颜色表示基因的表达水平;点大小代表在活化的 CD4+T 细胞群体中表达该基因的细胞分数。下图:来自 COVID-19 患者(COV)(n = 16),IAV 患者(IAV)(n = 4)和健康供体(Ctrl)(n = 3)的样本之间基因表达的差异。方框图中的每个点代表一个样品中激活的 CD4+T 细胞群体中某个基因的平均表达水平。使用学生 t 检验来检验差异的显著性。∗p <0.05,∗∗p <0.01,∗∗∗p <0.001。
(C)与(B)相似,显示了在 COVID-19 患者的活化 CD4+T 细胞中高表达的四个代表性基因。
另请参阅图S4。

参与宿主的免疫反应,其中包括多个关键转录因子STAT3,NFKB1,和REL,在从 IAV 例(各种细胞类型中上调图4A 和S4A)。NFKB1和REL编码 NF-κB异二聚体的活性亚基,NF-κB异二聚体是被IAV感染激活的标志性转录因子之一(Ludwig和Planz,2008年)。NF-κB的激活在调节促炎性先天免疫应答和适应性免疫应答中起关键作用。先前的研究表明,严重的IAV患者中STAT3信号传导的已知激活因子IL-6和IL-10显著增加(Yu等人,2011年)。与 COVID-19 患者和健康对照组相比,IAV 患者中STAT3 的表达升高,并且似乎与入院后的时间相关。此外,与其他组相比,IAV 患者的活化 CD4+T 细胞中RUNX3表达上调(图4B)。有人认为,RUNX3 诱导是 CD4+T 细胞获得细胞毒活性的关键步骤,而另一项研究表明,IAV 通过 NF-κB途径感染的RUNX3促进了气道上皮细胞的凋亡(Gan等,2015)。)。RUNX3 在 T 细胞介导的免疫反应中的作用尚待确定。几种前病毒宿主因素,例如CHD1,BCLAF1和PHF3有助于病毒感染,复制和免疫逃避也上调在 IAV 例(图4A 和S4A)(夏皮拉等人,2009年)。为了了解这些基因的响应随时间如何变化,作者将 IAV 患者的疾病过程分为两个阶段(表S3)。作者没有发现任何具有统计学意义的时间依赖性调节基因,尽管其中一些具有表达增加或减少的趋势(图S4B 和 S4C)。
对于 COVID-19 患者,STAT1 是响应干扰素而激活的主要转录因子,在激活的 CD4+T 细胞,细胞毒性 CD8+T 细胞,幼稚 T 细胞和 DC 中上调(图4A 和S4A)。活化的 CD4+T 细胞,细胞毒性 CD8+T 细胞,MAIT 细胞和 NK 细胞中的几种促炎因子(包括 TNF 和 TNFSF14)升高(图S4A 和 S4E),表明效应子功能和记忆细胞发育增强(Desai等。,2018)。IL6 的表达在任何患者或健康对照的 PBMC 中均未检出。作者在这些 COVID-19 患者的住院期间以及出院后测量了 IL-6 的血浆浓度(图S4D)。五分之二的患者入院时血浆 IL-6 高于正常范围(0.0–7.0 pg / mL)(Chen等,2020),住院期间恢复正常,恢复后保持稳定。大多数 COVID-19 患者的 IL-6 处于上升阶段,表明有活跃的炎症反应,这在 IAV 患者中也观察到(Yu等人,2011年)。具有严重症状的患者住院时的 IL-6 水平要高得多,恢复正常血浆 IL-6 水平所花费的时间更长,这与疾病的严重程度相对应。
IL-6 通过与 IL-6R 结合而发挥其功能。然后,IL-6-IL-6R 复合物与 GP130(IL-6 受体亚基β[IL6ST])结合,GP130是由几种相关细胞因子共享的常见信号转导链(Mihara等,2012)。的表达IL6R在活化的 CD4 升至+T 细胞,幼稚 T 细胞,和 COVID-19 的患者的 DCs 相比 IAV 患者和健康对照(图4C 和S4A)。同时, 与健康对照相比,COVID-19 患者和 IAV 患者在各种细胞类型中IL6ST 的表达上调。据报道,IL-6R 经常从活化的 T 细胞膜上脱落,于是它与 IL-6 的可溶性形式结合并以反式作用在表达 IL6ST 的细胞上。通过 IL6ST 进行的此类 IL-6 反信号可能显著促进 IL-6 的促炎特性(Wolf等人,2014年),强调了当前研究中观察到的IL6ST上调的潜在重要性。
根据临床观察,COVID-19 患者的炎症细胞因子(例如 IFN-α2,IL-7,IL-17和IL-10)的血浆浓度高于健康成年人(Huang等,2020)。在 COVID-19 患者,IAV 患者和健康对照中比较了活化 CD4+T 细胞,细胞毒性 CD8+T 细胞,MAIT 细胞和 NK 细胞中相应受体的表达水平。(图S4E)。与健康对照组相比,在 COVID-19 患者的这些细胞类型中,IFNAR1(干扰素α/β受体的α链)的上调是显著的。值得注意的是,在激活的CD4+T ,IL-7,IL-17和IL-27的受体亚基与IFNAR1一起显著升高COVID-19患者的T细胞与其他组相比,表明CD4+T细胞可能参与了对细胞因子的主要炎症反应。
最后,作者检查了它们表达的时间变化(图S4B 和 S4C)。尽管某些 IFN 反应性转录因子基因(例如STAT1和IRF3)随着时间的推移趋于降低表达水平,但由于患者数量有限,这些模式均无统计学意义。将来有更多患者和更多时间点的研究将解决这个问题。
讨论
新兴和再出现的病毒对人类健康构成持续威胁(高,2018)。新的 SARS-CoV-2 病毒感染会导致严重的肺部疾病和并发症,发病率和死亡率都很高。当前,没有针对这种致命性肺病的最佳治疗方法或有效药物。作者目前对宿主对 SARS-CoV-2 感染的免疫反应的理解是有限的,这使得设计急需的新型疗法变得困难。在这里,作者对来自 COVID-19 患者的 PBMC 进行了 scRNA-seq。总体而言,SARS-CoV-2 的感染对 PBMC 中免疫细胞的组成影响很小。在所有免疫细胞簇中,与健康对照组相比,所有五名 COVID-19 患者中浆细胞的百分比均显著增加。浆细胞的这种增加可以产生多种保护性中和抗体。这些对 SARS-CoV-2 的应答产生的 B 细胞来源的抗体对于预防急性呼吸道感染导致的死亡以及为免受未来感染引起的疾病和/或死亡提供持续保护至关重要。另外,目前鼓励包含来自康复患者的保护性中和抗体的血浆用于治疗重症患者。
在此过程中,病毒感染可诱导 IFN 发挥抗病毒功能并平衡病毒控制和免疫病理。在感染 SARS-CoV-2 的 ICU 患者中,血浆 IFNγ和TNF-α水平较高(Huang等,2020)。IFNα2和IFNγ在COVID-19中也与肺损伤密切相关(Liu等人,2020年)。此外,与健康对照组相比,仅在SARS-CoV-2感染的重症监护病房(ICU)患者中发现了共表达Th1细胞的IFNγ和粒细胞巨噬细胞集落刺激因子(GM-CSF)共表达(Zhou等,2020c) 。一致地,在作者的研究中,“ IFN-I 反应”丰富于 COVID-19 患者的不同 PBMC 亚群中。此外,ISG 的表达包括ISG15,IFI44L,与三个健康对照组相比,这些患者中的MX1和XAF1显著上调,表明 IFN 可能触发了强大的抗病毒功能(Perng和Lenschow,2018年)。此外,与轻度和健康患者相比,重症 COVID-19 患者对 IFN 和病毒感染的反应更强。这表明干扰素反应的强度既可以指示 COVID-19 疾病的严重程度,又可以将 COVID-19 患者与健康个体区分开。另外,IFN 应答的动力学表明个体差异,并表明针对 SARS-CoV-2 感染的 IFN 治疗的时机是关键的。
根据作者的结果,上调XAF1表达可能参与增加的 T 细胞凋亡 COVID-19 的患者中,与其它基因,包括配合IRF1,TP53,BCL2L11,和CASP3(Jeong等人,2018。;。Zou等人,2012)。一致地,在另一项 COVID-19 研究中,COVID-19 患者的TP53表达增加(Xiong等,2020)。除 XAF1 诱导的凋亡外,凋亡的外在途径,包括 TNF-α/ TNFR1和Fas / FasL途径(Elmore,2007年)),也被发现与 COVID-19 患者的不同细胞亚型有关。此外,在作者的研究中,据报道在严重的 COVID-19 病例中血浆 TNF-α升高(Chen等,2020;Huang等,2020),并且在本研究中 COVID-19 患者的 PBMC 中 TNF 的表达上调,证实了在 COVID-19 患者中 TNF-α分泌增加与TNF-α诱导的凋亡之间的相关性(Rath和Aggarwal,1999年)。XAF1 可以被 TNF-α和IFN诱导,并作为TNF-α诱导的细胞凋亡的替代途径(Strasszewski-Chavez等,2007)。XAF1 还被报道与 TNFSF10(TRAIL)共同促进登革热病毒诱导的细胞凋亡(Zhang等,2019)。综上所述,作者发现与 XAF1,TNF 和 Fas 途径相关的上调基因可能导致 COVID-19 患者的 T 细胞凋亡增加。
作者分析了细胞因子,细胞因子受体和转录因子的表达,这些因子在病毒感染的免疫反应中至关重要,并确定了与 IAV 患者不同的 COVID-19 患者的基因表达模式。细胞因子受体的上调与血清细胞因子水平的升高相一致,这可能会增强细胞因子介导的炎症反应。在 PBMC 中未检测到大多数细胞因子的表达这一事实可能表明血清细胞因子主要来自感染部位,即 COVID-19 的下呼吸道。作者发现大多数 COVID-19 患者的血浆 IL-6 浓度均高于正常水平,与其他研究一致(Chen等人,2020年)。在 IAV 患者中也有 IL-6 升高的报道(Yu等人,2011),它是急性呼吸应激综合症患者的免疫特征(Wang等人,2020c),与细胞因子释放综合症导致的死亡率有关。目前,已经批准了针对肺炎和 IL-6 升高的患者使用阻断 IL-6 或 IL-6R 的治疗方法],并且一项小型研究表明,针对 IL-6R 的单克隆抗体 Tocilizumab 可以缓解临床症状(Xu et al。,2020b)。作者的数据显示,在 COVID-19 患者中IL6R和IL6ST 的表达上调,并且作者建议IL6R和IL6ST 的表达增加 可能与升高的 IL-6 协同作用以诱导强烈的炎症反应,表明患者可能受益于 IL-6 或 IL-6R 拮抗剂治疗。
研究的局限性
该研究的解释存在重要局限性。
首先,因为在作者的研究中仅检查了有限的患者,尤其是对于 IAV(两名患者),所以作者需要通过更大的临床试验和/或进一步的研究来进一步验证在感染期间作者确定的 COVID-19 和 IAV 之间的差异。
第二,作者的研究集中于血液中 PBMC 的单细胞转录组。如果作者可以将作者的数据与病变部位(例如肺)的数据结合起来,作者的分析将更加系统化,因此结论将更加全面。
作者的研究在单细胞转录组水平上观察了与 IAV 患者相比,COVID-19 患者在疾病过程中免疫反应的动态格局。这些结果不仅代表了 COVID-19 临床过程中的免疫分子和细胞特征,而且还为诊断这种新疾病急需的生物标志物和治疗靶标提供了途径。
数据和代码可用性
原始和处理后的数据可在 CNGB 核苷酸序列档案库(CNSA:https: //db.cngb.org/cnsa)上获得,登录号为 CNP0001102。支持当前研究的代码可应要求从相应的作者处获得。
无监督群集
细胞聚类由 Seurat(v3.1)R 工具箱(Butler et al。,2018)(https://github.com/satijalab/seurat)。滤出少于 3 个细胞中表达的基因,排除检测到少于 300 个或超过 6000 个基因的细胞。为了处理批处理效果,分别对每个样本执行了“ NormalizeData”和“ FindVariableGene”功能(n = 23)。之后,使用“ FindIntergrationAnchors”和“ IntegrateData”函数将这 23 个批次集成在一起,并将 dims 参数设置为 20。然后,对集成数据集进行缩放并计算 PCA。首先使用 20 台 PC 来构建 SNN 网络,然后使用基于图的聚类方法 louvain 算法来识别分辨率设置为 1 的细胞簇。最后,使用 UMAP 来可视化二维空间中的聚类结果。为了进一步消除红细胞的影响,HBA1,HBA2,HBB,HBG1,HBG2,HBQ1,HBD,HBM,HBE1和HBZ),并使用上述相同方法再次执行聚类。
聚类标记分析和细胞类型注释
为了将每个簇注释为特定的免疫细胞类型,作者为免疫细胞选择了一些经典标记,并使用小提琴图(图1D)和 UMAP 特征图(图S1)来注释细胞类型。以下基因用于细胞类型注释:CD3G, CD4, CD8A, CD8B (T cells); KLRF1, XCL1 (NKs); MS4A1 (B cells); CD27 (Memory B cells) IGHG1, MZB1 (Plasma cells); CD68 (Monocytes); CD1C, LYZ (DCs); MKI67, TOP2A (Cycling cells); GZMA (Cytotoxic CD8+ T cells / NKs); CCR7, SELL (Naive T cells); PPBP (Megakaryocytes); CD34 (Stem cells).
差异表达基因(DEG)分析和 GO 富集
作者使用 Seurat R 软件包中的“ FindMarkers”功能进行了 DEG 分析。作者比较的细胞群分别作为 ident.1 和 ident.2 输入。计算所选细胞群之间基因平均表达水平的倍数变化。为了找到上调基因的功能(调整后的 p <0.01,Foldchange> 2),作者使用了功能 compareCluster(fun =“ enrichGO”,pvalueCutoff = 0.1,pAdjustMethod =“ BH”,OrgDb =“ org.Hs.eg.db ChIPseeker R 软件包(v.1.22.1)(Yu et al。,2015)。
下载:下载Acrobat PDF文件(5MB)
文件 S1。图 S1-S4。
下载:下载电子表格(12KB)
表 S1。入选受试者的临床数据,与图 1 相关。
下载:下载电子表格(10KB)
表 S2。临床实验室测试,与图 2 相关。
下载:下载电子表格(9KB)
表 S3。与图 3 相关的疾病进展阶段信息。
下载:下载Acrobat PDF文件(8MB)
文件 S2。文章加上补充信息。