- 微服务下功能权限与数据权限的设计与实现
nbsaas-boot
微服务java架构
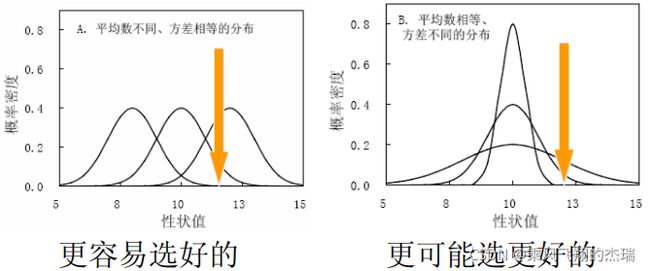
在微服务架构下,系统的功能权限和数据权限控制显得尤为重要。随着系统规模的扩大和微服务数量的增加,如何保证不同用户和服务之间的访问权限准确、细粒度地控制,成为设计安全策略的关键。本文将讨论如何在微服务体系中设计和实现功能权限与数据权限控制。1.功能权限与数据权限的定义功能权限:指用户或系统角色对特定功能的访问权限。通常是某个用户角色能否执行某个操作,比如查看订单、创建订单、修改用户资料等。数据权限:
- 在一台Ubuntu计算机上构建Hyperledger Fabric网络
落叶无声9
区块链超级账本Hyperledgerfabric区块链ubuntu构建hyperledgerfabric
在一台Ubuntu计算机上构建HyperledgerFabric网络Hyperledgerfabric是一个开源的区块链应用程序平台,为开发基于区块链的应用程序提供了一个起点。当我们提到HyperledgerFabric网络时,我们指的是使用HyperledgerFabric的正在运行的系统。即使只使用最少数量的组件,部署Fabric网络也不是一件容易的事。Fabric社区创建了一个名为Cello
- 【目标检测数据集】卡车数据集1073张VOC+YOLO格式
熬夜写代码的平头哥∰
目标检测YOLO人工智能
数据集格式:PascalVOC格式+YOLO格式(不包含分割路径的txt文件,仅仅包含jpg图片以及对应的VOC格式xml文件和yolo格式txt文件)图片数量(jpg文件个数):1073标注数量(xml文件个数):1073标注数量(txt文件个数):1073标注类别数:1标注类别名称:["truck"]每个类别标注的框数:truck框数=1120总框数:1120使用标注工具:labelImg标注
- 番茄西红柿叶子病害分类数据集12882张11类别
futureflsl
数据集分类数据挖掘人工智能
数据集类型:图像分类用,不可用于目标检测无标注文件数据集格式:仅仅包含jpg图片,每个类别文件夹下面存放着对应图片图片数量(jpg文件个数):12882分类类别数:11类别名称:["Bacterial_Spot_Bacteria","Early_Blight_Fungus","Healthy","Late_Blight_Water_Mold","Leaf_Mold_Fungus","Powdery
- 钢筋长度超限检测检数据集VOC+YOLO格式215张1类别
futureflsl
数据集YOLO深度学习机器学习
数据集格式:PascalVOC格式+YOLO格式(不包含分割路径的txt文件,仅仅包含jpg图片以及对应的VOC格式xml文件和yolo格式txt文件)图片数量(jpg文件个数):215标注数量(xml文件个数):215标注数量(txt文件个数):215标注类别数:1标注类别名称:["iron"]每个类别标注的框数:iron框数=215总框数:215使用标注工具:labelImg标注规则:对类别进
- 闲鱼鱼小铺怎么开通?鱼小铺开通需要哪些流程?
高省APP大九
闲鱼鱼小铺是平台推出的一个专业程度的店铺,与普通店铺相比会有更多的权益,比如说发布的商品数量从50增加到500;拥有专业的店铺数据看板与分析的功能,这对于专门在闲鱼做生意的用户来说是非常有帮助的,那么鱼小铺每个人都能开通吗?大家好,我是高省APP联合创始人蓓蓓导师,高省APP是2021年推出的电商导购平台,0投资,0风险、高省APP佣金更高,模式更好,终端用户不流失。【高省】是一个可省钱佣金高,能
- 目前哪里有卖高仿包包,推荐十个渠道已更新
富腕表之家
1、工厂购买,推荐微信:【76929666】目前买的人最多的渠道。2、某宝购买,价格较高,质量没有保障。3、拼夕夕,价格是便宜,但是质量不敢想象。4、专柜购买,数量较少,经常断货,价格也太高不好接受。5、批发市场购买,可遇不可求,一般生活在批发市场附近的,根本不用考虑在哪里买高仿包包分几个级别?在当今的包类市场中,广州作为一个知名的货源地,已经成为高仿包行业的一个重要标志。随着市场的需求增加,高仿
- 网络通信流程
记得开心一点啊
服务器网络运维
目录♫IP地址♫子网掩码♫MAC地址♫相关设备♫ARP寻址♫网络通信流程♫IP地址我们已经知道IP地址由网络号+主机号组成,根据IP地址的不同可以有5钟划分网络号和主机号的方案:其中,各类地址的表示范围是:分类范围适用网络网络数量主机最大连接数A类0.0.0.0~127.255.255.255大型网络12616777214【(2^24)-2】B类128.0.0.0~191.255.255.255中
- 5分钟说透AppStore审核原理,让你拥有上架新思路!
Q仔本人噢
在AppStore上架是越来越难了!相信非常多公司的技术人员都为此困扰,然而外包团队水平又层次不齐,容易遇坑,实在是内忧外患。是什么原因导致审核机制频繁调整?又是什么原因使得审核变得越发严格?那么接下来听小Q分解,马上给各位带来解答!首先看一下近一年的上下架的情况:近一年上架情况近一年下架情况通过数据我们发现越是马甲包产量权重高的分类里被下架的app数量越多,苹果此举可谓是上有政策,下有对策。通过
- SpringCloudAlibaba—Sentinel(限流)
菜鸟爪哇
前言:自己在学习过程的记录,借鉴别人文章,记录自己实现的步骤。借鉴文章:https://blog.csdn.net/u014494148/article/details/105484410Sentinel介绍Sentinel诞生于阿里巴巴,其主要目标是流量控制和服务熔断。Sentinel是通过限制并发线程的数量(即信号隔离)来减少不稳定资源的影响,而不是使用线程池,省去了线程切换的性能开销。当资源
- Day777--【这届年轻人很懒,“但在你面前,我可以多受些麻烦。”】
卖小妞的饼干
图片发自App中学时期,会因为朋友一句误会,手写三页信件解释来龙去脉,必须每天一起上学放学吃饭上洗手间,永远腻在一起才能彰显关系密切。而现在,相比每段关系都闹得轰轰烈烈,我们更喜欢顺其自然。懒得耗费大量时间去维护一段关系,懒得在与朋友产生矛盾后去辩解,懒得因为朋友做出背叛友谊的行为而愤怒诘问。不再把目光放在朋友的数量上,更在乎每一个还在身边的人是否饱含真心。我们都在期待着更舒服自然没有杂质的关系,
- vue+el-table 可输入表格使用上下键进行input框切换
以对_
vue学习记录vue.jsjavascript前端
使用上下键进行完工数量这一列的切换-->//键盘触发事件show(ev,index){letnewIndex;letinputAll=document.querySelectorAll('.table_inputinput');//向上=38if(ev.keyCode==38){if(index==0){//如果是第一行,回到最后一个newIndex=inputAll.length-1}elsei
- 分享十个渠道:高仿手表在哪里买最好的优选渠道
百奢之家
1、工厂购买,推荐靠谱手表商家微信:【15266277】目前买的人最多的渠道。2、某宝购买,价格较高,质量没有保障。3、拼夕夕,价格是便宜,但是质量不敢想象。4、专柜购买,数量较少,经常断货,价格也太高不好接受。5、批发市场购买,可遇不可求,一般生活在批发市场附近的,根本不用考虑在哪里买手表作为一件配饰品,不仅可以提升整体造型品味,还能展现个人的时尚态度。而对于很多人来说,高仿手表是一种经济实惠又
- mysql学习教程,从入门到精通,TOP 和MySQL LIMIT 子句(15)
知识分享小能手
大数据数据库MySQLmysql学习oracle数据库开发语言adb大数据
1、TOP和MySQLLIMIT子句内容在SQL中,不同的数据库系统对于限制查询结果的数量有不同的实现方式。TOP关键字主要用于SQLServer和Access数据库中,而LIMIT子句则主要用于MySQL、PostgreSQL(通过LIMIT/OFFSET语法)、SQLite等数据库中。下面将分别详细介绍这两个功能的语法、语句以及案例。1.1、TOP子句(SQLServer和Access)1.1
- 高性能javascript--算法和流程控制
海淀萌狗
-for,while和do-while性能相当-避免使用for-in循环,==除非遍历一个属性量未知的对象==es5:for-in遍历的对象便不局限于数组,还可以遍历对象。原因:for-in每次迭代操作会同时搜索实例或者原型属性,for-in循环的每次迭代都会产生更多开销,因此要比其他循环类型慢,一般速度为其他类型循环的1/7。因此,除非明确需要迭代一个属性数量未知的对象,否则应避免使用for-i
- [数据集][目标检测]汽车头部尾部检测数据集VOC+YOLO格式5319张3类别
FL1623863129
数据集目标检测汽车YOLO
数据集制作单位:未来自主研究中心(FIRC)版权单位:未来自主研究中心(FIRC)版权声明:数据集仅仅供个人使用,不得在未授权情况下挂淘宝、咸鱼等交易网站公开售卖,由此引发的法律责任需自行承担数据集格式:PascalVOC格式+YOLO格式(不包含分割路径的txt文件,仅仅包含jpg图片以及对应的VOC格式xml文件和yolo格式txt文件)图片数量(jpg文件个数):5319标注数量(xml文件
- Codeforces Round 972 (Div. 2) A-C 题解
AKDreamer_HeXY
Codeforces比赛题解c++算法动态规划数据结构贪心算法
本来以为B2难度会1900什么的,结果感觉1200还没有,先做的B1,后悔了QwQ关于我现场没切出C这件事……现场排名:A.SimplePalindrome题意构造一个长度为nnn的字符串,只包含aeiou五种字母,需要使得构造出来的字符串所包含的回文子序列数量最小思路当n≤5n\le5n≤5时,只要555个字母不重复出现都是最优情况当n>5n>5n>5时,可以证明:把相同字母放在一起是最优情况:
- 全视通智慧病房系统旧病房改造方案
2301_78035670
解决方案人工智能大数据
一、背景介绍在当今医疗技术日新月异的时代,智慧病房作为医院现代化建设的重要一环,正逐步从概念走向现实,深刻改变着患者的就医体验与医护人员的工作模式。智慧病房的改造背景,根植于医疗需求的日益增长、技术创新的不断推动以及对医疗质量与效率的不懈追求之中。二、医疗需求的深刻变革随着人口老龄化的加剧和慢性病患者数量的激增,医疗资源分配不均、医患沟通不畅、患者照护效率低下等问题日益凸显。传统的病房管理模式已难
- 2022-07-06
榜一大哥啊
非洲猪瘟检测流程要点1、进入实验室按照要求穿好装备进入实验室,病原稀释及制备,将实验用假阳性按照倍数稀释,最高稀释到一万倍。所有操作流程都在生物安全柜进行,按照流程进行编号,编写检测编号。在每个实验室都要将白大褂以及手套进行更换。2、到试剂准备区进行试剂准备,按照样品数量加阴阳对照进行配备,该项目在超净工作台进行。将制备好的试剂放入传递窗,进入核酸提取环节。3、核酸提取区,进行核酸提纯,用磁吸法核
- app推广一手资源在哪里找?盘点2024年必备的八大app拉新渠道
U客直谈APP
在2024年即将来临之际,还是有许多小伙伴表示不知道app推广一手资源在哪里找,又要从哪里去了解各个资源渠道的不同特性。好消息来啦,本篇文章就将带大家盘点24年必备的八大app拉新渠道,全文干货,拆解分析点评一步到位,还不快快收藏起来~1.app推广一手资源来源:U客直谈U客直谈是一个资源对接平台,专注于为推广人员提供海量丰富的app拉新任务。其具有数量丰富且类型多样的app拉新任务,使得U客直谈
- 跟着黑马学mysql(5)
小杜不吃糖
mysql数据库
17.DQL-聚合函数DQL-聚合函数介绍将一列数据作为一个整体,进行纵向计算。常见聚合函数函数功能count统计数量max最大值min最小值avg平均值sum求和语法SELECT聚合函数(字段列表)FROM表名;注意:所有的null值不参与聚合函数的运算18.DQL-分组查询语法SELECT字段列表FROM表名[WHERE条件]GROUPBY分组字段名[HAVING分组后的过滤条件];where
- Github 2024-09-12 Go开源项目日报Top10
老孙正经胡说
githubgolang开源Github趋势分析开源项目PythonGolang
根据GithubTrendings的统计,今日(2024-09-12统计)共有10个项目上榜。根据开发语言中项目的数量,汇总情况如下:开发语言项目数量Go项目10C项目1Terraform:基础设施即代码的开源工具创建周期:3626天开发语言:Go协议类型:OtherStar数量:40393个Fork数量:9397次关注人数:40393人贡献人数:358人OpenIssues数量:1943个Git
- 两岁娃不会数数?正常!帮娃跨越数字关,点数识数量数对应都要会
说书人熊二娘
前几天,我遇到一个很久不见的年长朋友。他向我吐槽自己的孙子不会数数。他对我说自己有一天拿出了道具,让小孙子学数。两岁多的小孙子看了看那些道具,对他说:“爷爷,我们今天还是算了吧。”两岁多的孩子学数的确是早了一点。孩子的认知水平没有到,学起东西来就会异常困难。等过了一段时间,他认知和理解力达到了学数的程度,分分钟就能搞定这些问题。我儿子3岁7个月,最近突然能够数到100,玩起百数板也非常开心。我偶尔
- Golang Channel
PandaSkr
golang
Channel解析1.Channel源码分析1.1Channel数据结构typehchanstruct{qcountuint//channel的元素数量dataqsizuint//channel循环队列长度bufunsafe.Pointer//指向循环队列的指针elemsizeuint16//元素大小closeduint32//channel是否关闭0-未关闭elemtype*_type//元素类
- 2018-08-11
daring婧
贩卖毒品是法律严厉打击的行为,却依然有人铤而走险、以身试法,但最终得到的只会是法律的惩罚。贩卖毒品谋取小利,不知悔改“二进宫”黄某曾于2016年12月29日因贩卖毒品罪被钦州市钦南区人民法院判处有期徒刑十个月,缓刑一年。但得到缓刑机会的黄某却不以为然,认为只要贩卖毒品的数量不大,就不会被抓进看守所,依旧继续贩卖着毒品。2018年4月11日,黄某在钦州市钦南区四马路将1小包毒品海洛因(净重0.1克)
- 算法笔试-编程练习-好题-04
Glen 997
大厂校招-编程集训算法大厂笔试动态规划堆盒子
题目:堆盒子礼盒大小不同,我们玩堆盒子的游戏,怎么堆盒子使得堆出的高度最高,每个礼盒的大小由长、宽、高表示,堆盒子的时候要求下面的盒子长、宽、高都必须大于上面的盒子,不包含等于。高度为堆出的礼盒的所有高度的总和。输入描述输入的第一行是礼盒的个数N,接下来输入N行,每行表示每个礼盒的长、宽、高。礼盒的数量不超过1000个,每个盒子的长、宽、高取值范围为1~10。输出描述输出一行,输出能堆出盒子的最高
- php服务器状态监测,PHP网站状态在线监控源码
傲雪吟霜白如冰
php服务器状态监测
在网上找的一个在线监控源码,比较简单,但是功能也略有不足,例如如果网站挂了就按照监控频率一直发邮件提示,网站恢复之后不会发邮件通知;不能直接填写要监控的网址或者某一页面,适合监控大量的网站或者vps、服务器。最新使用感受:没有发送邮件限制,我设置监控频率是一个小时,一天没管就把邮箱塞满了。如果服务器恢复的话不会专门发邮件提醒。如果只是监控几个站的话有很多免费的监控网站可以使用,只是使用数量上有限制
- JVM 调优篇7 调优案例1-堆空间的优化解决
健康平安的活着
jvm调优jvm
一jvm优化1.1优化实施步骤*1)减少使用全局变量和大对象;2)调整新生代的大小到最合适;3)设置老年代的大小为最合适;4)选择合适的GC收集器;1.2关于GC优化原则多数的Java应用不需要在服务器上进行GC优化;多数导致GC问题的Java应用,都不是因为我们参数设置错误,而是代码问题;在应用上线之前,先考虑将机器的JVM参数设置到最优(最适合);减少创建对象的数量;减少使用全局变量和大对象;
- 图片中的上采样,下采样和通道融合(up-sample, down-sample, channel confusion)
迪三
#图像处理_PyTorch计算机视觉深度学习人工智能
前言以conv2d为例(即图片),Pytorch中输入的数据格式为tensor,格式为:[N,C,W,H,W]第一维N.代表图片个数,类似一个batch里面有N张图片第二维C.代表通道数,在模型中输入如果为彩色,常用RGB三色图,那么就是3维,即C=3。如果是黑白的,即灰度图,那么只有一个通道,即C=1第三维H.代表图片的高度,H的数量是图片像素的列数第四维W.代表图片的宽度,W的数量是图片像素的
- 自己看---华为od--构成正方形的数量
我狠狠地刷刷刷刷刷
华为od算法
题目描述输入N个互不相同的二维整数坐标,求这N个坐标可以构成的正方形数量。[内积为零的的两个向量垂直]输入描述第一行输入为N,N代表坐标数量,N为正整数。N<=100之后的N行输入为坐标xy以空格分隔,x,y为整数,-10<=x,y<=10输出描述输出可以构成的正方形数量。用例1输入3132431输出0用例2输入40012312-1输出1N=int(input())coords=[input()f
- 312个免费高速HTTP代理IP(能隐藏自己真实IP地址)
yangshangchuan
高速免费superwordHTTP代理
124.88.67.20:843
190.36.223.93:8080
117.147.221.38:8123
122.228.92.103:3128
183.247.211.159:8123
124.88.67.35:81
112.18.51.167:8123
218.28.96.39:3128
49.94.160.198:3128
183.20
- pull解析和json编码
百合不是茶
androidpull解析json
n.json文件:
[{name:java,lan:c++,age:17},{name:android,lan:java,age:8}]
pull.xml文件
<?xml version="1.0" encoding="utf-8"?>
<stu>
<name>java
- [能源与矿产]石油与地球生态系统
comsci
能源
按照苏联的科学界的说法,石油并非是远古的生物残骸的演变产物,而是一种可以由某些特殊地质结构和物理条件生产出来的东西,也就是说,石油是可以自增长的....
那么我们做一个猜想: 石油好像是地球的体液,我们地球具有自动产生石油的某种机制,只要我们不过量开采石油,并保护好
- 类与对象浅谈
沐刃青蛟
java基础
类,字面理解,便是同一种事物的总称,比如人类,是对世界上所有人的一个总称。而对象,便是类的具体化,实例化,是一个具体事物,比如张飞这个人,就是人类的一个对象。但要注意的是:张飞这个人是对象,而不是张飞,张飞只是他这个人的名字,是他的属性而已。而一个类中包含了属性和方法这两兄弟,他们分别用来描述对象的行为和性质(感觉应该是
- 新站开始被收录后,我们应该做什么?
IT独行者
PHPseo
新站开始被收录后,我们应该做什么?
百度终于开始收录自己的网站了,作为站长,你是不是觉得那一刻很有成就感呢,同时,你是不是又很茫然,不知道下一步该做什么了?至少我当初就是这样,在这里和大家一份分享一下新站收录后,我们要做哪些工作。
至于如何让百度快速收录自己的网站,可以参考我之前的帖子《新站让百
- oracle 连接碰到的问题
文强chu
oracle
Unable to find a java Virtual Machine--安装64位版Oracle11gR2后无法启动SQLDeveloper的解决方案
作者:草根IT网 来源:未知 人气:813标签:
导读:安装64位版Oracle11gR2后发现启动SQLDeveloper时弹出配置java.exe的路径,找到Oracle自带java.exe后产生的路径“C:\app\用户名\prod
- Swing中按ctrl键同时移动鼠标拖动组件(类中多借口共享同一数据)
小桔子
java继承swing接口监听
都知道java中类只能单继承,但可以实现多个接口,但我发现实现多个接口之后,多个接口却不能共享同一个数据,应用开发中想实现:当用户按着ctrl键时,可以用鼠标点击拖动组件,比如说文本框。
编写一个监听实现KeyListener,NouseListener,MouseMotionListener三个接口,重写方法。定义一个全局变量boolea
- linux常用的命令
aichenglong
linux常用命令
1 startx切换到图形化界面
2 man命令:查看帮助信息
man 需要查看的命令,man命令提供了大量的帮助信息,一般可以分成4个部分
name:对命令的简单说明
synopsis:命令的使用格式说明
description:命令的详细说明信息
options:命令的各项说明
3 date:显示时间
语法:date [OPTION]... [+FORMAT]
- eclipse内存优化
AILIKES
javaeclipsejvmjdk
一 基本说明 在JVM中,总体上分2块内存区,默认空余堆内存小于 40%时,JVM就会增大堆直到-Xmx的最大限制;空余堆内存大于70%时,JVM会减少堆直到-Xms的最小限制。 1)堆内存(Heap memory):堆是运行时数据区域,所有类实例和数组的内存均从此处分配,是Java代码可及的内存,是留给开发人
- 关键字的使用探讨
百合不是茶
关键字
//关键字的使用探讨/*访问关键词private 只能在本类中访问public 只能在本工程中访问protected 只能在包中和子类中访问默认的 只能在包中访问*//*final 类 方法 变量 final 类 不能被继承 final 方法 不能被子类覆盖,但可以继承 final 变量 只能有一次赋值,赋值后不能改变 final 不能用来修饰构造方法*///this()
- JS中定义对象的几种方式
bijian1013
js
1. 基于已有对象扩充其对象和方法(只适合于临时的生成一个对象):
<html>
<head>
<title>基于已有对象扩充其对象和方法(只适合于临时的生成一个对象)</title>
</head>
<script>
var obj = new Object();
- 表驱动法实例
bijian1013
java表驱动法TDD
获得月的天数是典型的直接访问驱动表方式的实例,下面我们来展示一下:
MonthDaysTest.java
package com.study.test;
import org.junit.Assert;
import org.junit.Test;
import com.study.MonthDays;
public class MonthDaysTest {
@T
- LInux启停重启常用服务器的脚本
bit1129
linux
启动,停止和重启常用服务器的Bash脚本,对于每个服务器,需要根据实际的安装路径做相应的修改
#! /bin/bash
Servers=(Apache2, Nginx, Resin, Tomcat, Couchbase, SVN, ActiveMQ, Mongo);
Ops=(Start, Stop, Restart);
currentDir=$(pwd);
echo
- 【HBase六】REST操作HBase
bit1129
hbase
HBase提供了REST风格的服务方便查看HBase集群的信息,以及执行增删改查操作
1. 启动和停止HBase REST 服务 1.1 启动REST服务
前台启动(默认端口号8080)
[hadoop@hadoop bin]$ ./hbase rest start
后台启动
hbase-daemon.sh start rest
启动时指定
- 大话zabbix 3.0设计假设
ronin47
What’s new in Zabbix 2.0?
去年开始使用Zabbix的时候,是1.8.X的版本,今年Zabbix已经跨入了2.0的时代。看了2.0的release notes,和performance相关的有下面几个:
:: Performance improvements::Trigger related da
- http错误码大全
byalias
http协议javaweb
响应码由三位十进制数字组成,它们出现在由HTTP服务器发送的响应的第一行。
响应码分五种类型,由它们的第一位数字表示:
1)1xx:信息,请求收到,继续处理
2)2xx:成功,行为被成功地接受、理解和采纳
3)3xx:重定向,为了完成请求,必须进一步执行的动作
4)4xx:客户端错误,请求包含语法错误或者请求无法实现
5)5xx:服务器错误,服务器不能实现一种明显无效的请求
- J2EE设计模式-Intercepting Filter
bylijinnan
java设计模式数据结构
Intercepting Filter类似于职责链模式
有两种实现
其中一种是Filter之间没有联系,全部Filter都存放在FilterChain中,由FilterChain来有序或无序地把把所有Filter调用一遍。没有用到链表这种数据结构。示例如下:
package com.ljn.filter.custom;
import java.util.ArrayList;
- 修改jboss端口
chicony
jboss
修改jboss端口
%JBOSS_HOME%\server\{服务实例名}\conf\bindingservice.beans\META-INF\bindings-jboss-beans.xml
中找到
<!-- The ports-default bindings are obtained by taking the base bindin
- c++ 用类模版实现数组类
CrazyMizzz
C++
最近c++学到数组类,写了代码将他实现,基本具有vector类的功能
#include<iostream>
#include<string>
#include<cassert>
using namespace std;
template<class T>
class Array
{
public:
//构造函数
- hadoop dfs.datanode.du.reserved 预留空间配置方法
daizj
hadoop预留空间
对于datanode配置预留空间的方法 为:在hdfs-site.xml添加如下配置
<property>
<name>dfs.datanode.du.reserved</name>
<value>10737418240</value>
- mysql远程访问的设置
dcj3sjt126com
mysql防火墙
第一步: 激活网络设置 你需要编辑mysql配置文件my.cnf. 通常状况,my.cnf放置于在以下目录: /etc/mysql/my.cnf (Debian linux) /etc/my.cnf (Red Hat Linux/Fedora Linux) /var/db/mysql/my.cnf (FreeBSD) 然后用vi编辑my.cnf,修改内容从以下行: [mysqld] 你所需要: 1
- ios 使用特定的popToViewController返回到相应的Controller
dcj3sjt126com
controller
1、取navigationCtroller中的Controllers
NSArray * ctrlArray = self.navigationController.viewControllers;
2、取出后,执行,
[self.navigationController popToViewController:[ctrlArray objectAtIndex:0] animated:YES
- Linux正则表达式和通配符的区别
eksliang
正则表达式通配符和正则表达式的区别通配符
转载请出自出处:http://eksliang.iteye.com/blog/1976579
首先得明白二者是截然不同的
通配符只能用在shell命令中,用来处理字符串的的匹配。
判断一个命令是否为bash shell(linux 默认的shell)的内置命令
type -t commad
返回结果含义
file 表示为外部命令
alias 表示该
- Ubuntu Mysql Install and CONF
gengzg
Install
http://www.navicat.com.cn/download/navicat-for-mysql
Step1: 下载Navicat ,网址:http://www.navicat.com/en/download/download.html
Step2:进入下载目录,解压压缩包:tar -zxvf navicat11_mysql_en.tar.gz
- 批处理,删除文件bat
huqiji
windowsdos
@echo off
::演示:删除指定路径下指定天数之前(以文件名中包含的日期字符串为准)的文件。
::如果演示结果无误,把del前面的echo去掉,即可实现真正删除。
::本例假设文件名中包含的日期字符串(比如:bak-2009-12-25.log)
rem 指定待删除文件的存放路径
set SrcDir=C:/Test/BatHome
rem 指定天数
set DaysAgo=1
- 跨浏览器兼容的HTML5视频音频播放器
天梯梦
html5
HTML5的video和audio标签是用来在网页中加入视频和音频的标签,在支持html5的浏览器中不需要预先加载Adobe Flash浏览器插件就能轻松快速的播放视频和音频文件。而html5media.js可以在不支持html5的浏览器上使video和audio标签生效。 How to enable <video> and <audio> tags in
- Bundle自定义数据传递
hm4123660
androidSerializable自定义数据传递BundleParcelable
我们都知道Bundle可能过put****()方法添加各种基本类型的数据,Intent也可以通过putExtras(Bundle)将数据添加进去,然后通过startActivity()跳到下一下Activity的时候就把数据也传到下一个Activity了。如传递一个字符串到下一个Activity
把数据放到Intent
- C#:异步编程和线程的使用(.NET 4.5 )
powertoolsteam
.net线程C#异步编程
异步编程和线程处理是并发或并行编程非常重要的功能特征。为了实现异步编程,可使用线程也可以不用。将异步与线程同时讲,将有助于我们更好的理解它们的特征。
本文中涉及关键知识点
1. 异步编程
2. 线程的使用
3. 基于任务的异步模式
4. 并行编程
5. 总结
异步编程
什么是异步操作?异步操作是指某些操作能够独立运行,不依赖主流程或主其他处理流程。通常情况下,C#程序
- spark 查看 job history 日志
Stark_Summer
日志sparkhistoryjob
SPARK_HOME/conf 下:
spark-defaults.conf 增加如下内容
spark.eventLog.enabled true spark.eventLog.dir hdfs://master:8020/var/log/spark spark.eventLog.compress true
spark-env.sh 增加如下内容
export SP
- SSH框架搭建
wangxiukai2015eye
springHibernatestruts
MyEclipse搭建SSH框架 Struts Spring Hibernate
1、new一个web project。
2、右键项目,为项目添加Struts支持。
选择Struts2 Core Libraries -<MyEclipes-Library>
点击Finish。src目录下多了struts