摘要
背景
Wilson病(WD)是由编码铜转运蛋白的ATP7B突变引起的常染色体隐性遗传疾病。随后的铜积累导致涉及肝,神经和精神症状的可变WD临床表型,没有明确的基因型 - 表型相关性。本研究的目的是分析来自WD患者的肝脏和血液中全基因组水平的DNA甲基化改变,以研究与WD诊断和表型相关的表观基因组改变。我们使用全基因组亚硫酸氢盐测序(WGBS)来检查WD受试者的不同群组,以确定DNA甲基化是否可以区分患者与健康受试者和患有其他肝脏疾病的受试者并区分不同的WD表型。
结果
肝脏中的WGBS分析鉴定了969个高甲基化区域和871个低甲基化差异甲基化区域(DMR),特异性识别WD患者,包括具有全基因组显着性的18个区域。WD特异性肝脏DMR与富含叶酸和脂质代谢功能以及急性炎症反应的基因相关,并且可以区分WD患者的早期纤维化。功能注释显示WD-高甲基化肝脏DMR富含肝脏特异性增强子,侧翼活性肝脏启动子和肝脏发育转录因子的结合位点,包括肝细胞核因子4α(HNF4A),维甲酸X受体α(RXRA),Forkhead方框A1(FOXA1)和FOXA2。还鉴定了与WD发育相关的DMR,包括具有全基因组显着性的15个。然而,肝脏中的WD DMR与肝细胞类型比例的大规模变化无关。在血液分化的WD患者中检测到来自健康和疾病对照受试者的DMR,并区分具有肝和神经性WD表现的患者。WD表型DMR对应于富含精神衰退,异常B细胞生理学功能的基因,以及作为多梳抑制复合物1(PRC1)的成员。在小验证队列中测试的与WD表型相关的44个DMR具有0.9的预测值。
结论
我们确定了WD的疾病机制相关的表观基因组特征,揭示了对这种复杂的单基因疾病的潜在生物标志物和治疗方法的新见解。
电子辅助材料
关键词:Wilson病,表观遗传学,铜,DNA甲基化,全基因组亚硫酸氢盐测序,染色质,肝脏,血液,生物标志物,增强子
背景
Wilson病(WD)是一种常染色体隐性疾病,由铜的积聚主要在肝脏和大脑中引起,这是由于突变影响铜转运基因,ATPase铜转运β(ATP7B)。在健康个体中,ATP7B有助于铜贩卖肝细胞内和所需的铜排泄到胆道[ 1,2 ]。然而,虽然现在更好地理解遗传基础,但由于对深层致病机制的不完全理解,缺乏金标准诊断测试以及最终有限的治疗选择,WD仍然代表临床上具有挑战性且往往未被认识的病症 [3]]。临床表现包括肝脏,神经和精神病表现。DNA序列突变与临床表现之间缺乏明确的相关性可归因于超过500种突变的存在,这些突变共同损害ATP7B铜转运蛋白活性,并且可能存在伴随的修饰基因。最引人注目的证据表明,遗传突变因素不谈影响表型从描述受WD单卵双生子但呈现不同的表型[多个病例报告衍生4,5 ]。
DNA甲基化是一种可逆的表观遗传修饰,受遗传和非遗传因素如营养和毒素暴露的影响,并作用于遗传和环境之间的界面。营养因素影响甲硫氨酸代谢和甲基对DNA和组蛋白甲基化的全球可用性。的WD变化显示甲硫氨酸代谢[动物模型6,7 ]和基因转录物水平响应于膳食规定甲基供体[的8]。特别是,已知铜会干扰S-腺苷同型半胱氨酸水解酶(SAHH)的表达和活性,而后者又会影响S-adenosylhomocysteine(SAH)水平和DNA甲基转移酶活性和表达[ 9 ]。
通过转录因子介导的DNA甲基转移酶和去甲基化酶的募集,可以在个别基因座处改变DNA甲基化,从而影响未来的转录因子结合效率[ 10 ]。基因启动子的甲基化与基因表达呈负相关,而基因体甲基化则呈正相关。在患有常见肝病的患者中,已经在血液和肝脏中观察到基因特异性DNA甲基化模式的变化,并且已经与肝病严重程度的不同阶段相关[ 11 ]。虽然肝脏和大脑是WD中受影响最大的组织,但血浆中的铜也会增加[ 12]。增加的体液免疫应答,降低的细胞介导的免疫应答,并在伽马Δ+ T细胞的增加已经在WD患者的血液中发现,这表明肝脏和血液可以具有特性甲基化的签名在WD [ 13,14 ] 。
有强有力的证据支持动物模型中WD发病机制中甲硫氨酸代谢的改变,以及间接证据支持相同机制在人类发病和进展中的作用[ 15 ]。在本研究中,我们探讨了这样的假设:肝脏和血液中的DNA甲基化模式可以区分健康受试者和受其他肝脏疾病影响的受试者(包括非酒精性脂肪性肝病(NAFLD))的各种表型表现的WD患者。 )和原发性硬化性胆管炎(PSC),最终目标是确定对致病机制,诊断标志物和治疗目标的新见解。
结果
肝脏样本的临床特征
共有21个肝脏样本可用于甲基化组分析,其中6个来自正常肝脏组织学的减肥手术的健康对照(HC),5个来自NAFLD(疾病对照,DC)的受试者,10个来自WD患者,包括来自肝硬化和急性肝衰竭患者的5例经皮肝活组织检查和5例来自外植肝脏的活检(附加文件1:表S1,附加文件2:表S2)。所有受试者的中位年龄为44岁(范围22-72,三组之间没有差异)。与WD患者相比,HC具有更高的BMI,因为样本来自减肥手术患者。正如预期的那样,与其他受试者相比,WD患者呈现出显着更晚期的纤维化阶段。
肝脏中的DMR将WD患者与健康和疾病对照区分开来
为了鉴定肝脏中WD特异性的DNA甲基化变化,来自WD(n = 10),HC(n = 6)和DC(n = 5)受试者的样品通过WGBS分析每组之间的DMR。鉴定了969个高甲基化区域和871个低甲基化区域,其特异性地区分WD与HC和DC受试者,但不是来自HC受试者的DC(图 1a)。在这些WD特异性DMR中,18个通过FWER达到全基因组显着性(表 1)。这18个DMR附近的基因包括那些在肝脏发育中具有已知功能的基因(BST1,FOXA1和VTN)和转录调控(FOXA1,MAFB,MN1,NACC2,ZNF689和ZNF785)。WD特异性肝脏DMR的基因本体分析显示,高甲基化DMR附近的基因富含与急性炎症反应,脂质分解代谢和叶酸代谢相关的功能(附加文件1:图S1)。相反,低甲基化DMR附近的基因富含与体液免疫应答,脂肪酸转运和糖酵解调节相关的功能。

)
图。1
肝脏DMR区分WD患者与对照组。鉴定了 WGBS衍生的DMR,其区分WD与HC和DC,但不区分来自HC的DC(WD n = 10,HC n = 6,DC n = 5)。高甲基化的DMR在WD中具有较高的甲基化,而低甲基化的DMR在WD中具有较低的甲基化。b使用WD特异性肝脏DMR中甲基化的HC,WD和DC热图。绘制每个样品相对于每个DMR的平均甲基化的甲基化百分比。C在WD特异性肝脏DMR中使用甲基化的主成分分析。每个点的大小表示患者的纤维化阶段。省略号显示95%的置信区间。对于这个和所有后续数字:DMR,差异甲基化区域; WD,Wilson病; WGBS,全基因组亚硫酸氢盐测序; HC,健康控制; DC,疾病控制
表格1
WD特异性肝脏DMR具有全基因组显着性和相关基因
| CHR | 开始 | 结束 | 化CpG | 甲基化差异(%) | FWER | 基因 | 到TSS的距离(kb) | 位置 |
|---|---|---|---|---|---|---|---|---|
| CHR21 | 31557558 | 31560049 | 156 | - 37 | <0.001 | Tiam 1 mRNA | 0.0 | TSS |
| LOC150051 | 0.0 | TSS | ||||||
| chr12 | 3199472 | 3201083 | 96 | - 35 | 0.004 | TSPAN9 | 122.1 | 内含子 |
| PRMT8 | - 180.3 | 上游 | ||||||
| CHR 9, | 136050881 | 136052939 | 57 | - 35 | 0.008 | NACC2 | 42.3 | 内含子 |
| UBAC1 | - 89.5 | 上游 | ||||||
| chr22 | 27797270 | 27798795 | 87 | 27 | 0.018 | MN1 | 2.7 | 外显子 |
| LOC100507657 | 486.6 | 下游 | ||||||
| CHR 15 | 99105435 | 99106614 | 93 | - 13 | 0.020 | SYNM | 0.4 | 外显子 |
| LRRC28 | - 144.7 | 上游 | ||||||
| CHR 18 | 22171324 | 22172395 | 66 | - 27 | 0.023 | GATA6 - AS1 | - 2.4 | 上游 |
| CHR21 | 44456208 | 44457837 | 56 | 29 | 0.023 | LRRC3 | 0.7 | 外显子 |
| LRRC3 - AS1 | - 0.9 | 上游 | ||||||
| chr20 | 40691316 | 40693007 | 60 | - 28 | 0.025 | MAFB | - 2.1 | 上游 |
| CHR 9, | 137461602 | 137462399 | 37 | - 28 | 0.028 | NSMF | - 2.3 | 上游 |
| CHR 9, | 136418051 | 136418847 | 29 | 32 | 0.031 | SDCCAG3 | - 7.4 | 上游 |
| INPP5E | 21.0 | 下游 | ||||||
| CHR11 | 66855700 | 66857160 | 56 | - 32 | 0.032 | LRFN4 | - 0.2 | 上游 |
| chr22 | 39994424 | 39995559 | 72 | - 24 | 0.035 | FAM83F | 0.0 | TSS |
| CHR 4 | 15702770 | 15703332 | 32 | - 28 | 0.038 | BST1 | 0.0 | TSS |
| chr14 | 37597362 | 37599120 | 65 | - 17 | 0.038 | FOXA1 | - 2.2 | 上游 |
| chr16 | 30604376 | 30605260 | 48 | 29 | 0.043 | ZNF689 | 5.5 | 外显子 |
| ZNF785 | - 18.6 | 上游 | ||||||
| chr17 | 28371994 | 28372583 | 59 | - 20 | 0.044 | SARM1 | 0.3 | 外显子 |
| VTN | - 1.6 | 上游 | ||||||
| CHR 9, | 121772727 | 121773248 | 32 | 23 | 0.048 | DAB2IP | 205.6 | 外显子 |
| TTLL11 | 320.4 | 下游 | ||||||
| CHR5 | 10649279 | 10650004 | 64 | 18 | 0.049 | ANKRD33B | 85.0 | 外显子 |
| DAP | 111.3 | 下游 |
FWER,家庭智能错误率
大多数WD受试者可从HC和DC受试者中清楚地区分甲基化水平的WD-肝特异性的DMR内的基础上(图 1 B,C)。这些DMR中的甲基化与性别,年龄,BMI,炎症或脂肪变性无关(附加文件1:图S2,附加文件2:表S3)。相反,在WD特定的DMRS整体,由第一主成分表示的甲基化,用纤维化(相关联的p = 2.5E-4),其也可以在WD的样品(图中观察到的异质性 1个 C)。单独地,23个DMR(1%的WD特异性DMR)与纤维化相关(Bonferroni调整后p <0.05,附加文件2):表S3)。此外,63例(3%)WD特异性DMR与先前在NAFLD受试者的纤维化甲基化研究中发现的DMR重叠[ 16 ]。然而,大多数个体WD特异性DMR与纤维化无显着相关性。
为了研究纤维化发病前肝脏中WD特异性DNA甲基化的变化,将早期(0-1期)WD患者(n = 3)与HC(n = 6)和早期DC(n = 4)科目。鉴定了124个高甲基化区域和70个低甲基化区域,区分早期WD患者,其与人口统计学协变量无关(附加文件1:图S3,附加文件2:表S4)。其中76个早期区域和167个相关基因与从所有WD患者中鉴定的WD特异性肝脏DMR重叠,显着富集(Hyper:p = 5.2E-82,Hypo:p = 1.4E-31)。此外,在WD的tx-j小鼠模型中,基于人WD差异甲基化选择的10个基因与病理早期的肝脏中的对照相比显着差异表达(q <0.05,附加文件1:表S5)。这些基因的GATA6,HDAC5,Pmpca,Pnpla7和Tspan9上调,而FOXA1,MAFB,Nacc2,PCX和的Vtn被下调了。这些结果一起表明,在人WD肝脏中检测到的甲基化差异具有功能性后果,并且与早期发病机制相关,而不是晚期纤维化的结果。
WD特异性高甲基化肝脏DMR富含肝脏增强剂和侧翼活性肝脏启动子
为了对从所有样本中鉴定的WD特异性肝脏DMR进行功能性注释,将Roadmap Epigenomics Project [ 17 ]中127种细胞和组织类型的组蛋白修饰ChIP-seq峰和染色质状态预测与DMR染色体位点进行比较(图 2))。高甲基化WD特异性DMR高度显着富集肝脏H3K4me1和H3K4me3组蛋白修饰标记分别重叠87%和50%的高甲基化DMR(H3K4me1优势比= 28.6,错误发现率(FDR)q <1.0E-319; H3K4me3赔率比率= 8.8,FDR q = 7.6E-210)。相反,低甲基化的WD特异性DMR在许多组织类型中富含H3K4me1和H3K4me3标记。接下来,WD特异性肝脏DMR与ChromHMM染色质状态预测重叠,后者使用组蛋白修饰ChIP-seq数据将基因组分成15个功能状态。相比在所有的组织背景中增强子(ENH,黄色)和区域侧翼活性转录起始位点(TssAFlnk,橙色)组合WD特定的DMRS显示显著富集(图 2 b)中,与这些功能区的一个显着显着富集(图 2 C)。高甲基化WD特异性DMR也分别在肝脏增强子和活性转录起始位点侧翼区域特异性富集(附加文件1):图S4; Enh比值比= 11.6,FDR q = 5.5E-278; TssAFlnk比值比= 10.1,FDR q = 3.7E-192)。转录区侧翼区域也显着富集,但仅构成一小部分高甲基化DMR。

)
WD特异性高甲基化肝脏DMR富含肝脏增强剂和侧翼活性肝脏启动子。使用LOLA 将 WD肝脏DMR与来自表观基因组路线图的组蛋白修饰ChIP-seq峰重叠,并对所有组织绘制比值比。b,c WD肝脏DMR与使用LOLA的表观基因组路线图中的染色质状态重叠,并且将每个状态重叠的DMR和背景区域的百分比绘制为b所有组织的平均重叠或肝脏的c重叠
肝脏中WD特异性高甲基化DMR富含肝相关转录因子结合位点
为了确定WD特异性肝脏DMR的甲基化差异与转录因子结合的潜在关联,DMR与来自ENCODE的肝脏中的转录因子ChIP-seq峰重叠(图 3a)并评估已知转录因子结合位点序列的富集。基序(图 3 b)中。高甲基化的DMR富含HNF4A,RXRA,FOXA1和FOXA2的肝结合位点和序列基序,而低甲基化的DMR没有富集这些因子(附加文件1:图S5)。当高甲基化DMR与HNF4A,RXRA,FOXA1和FOXA2的重叠结合位点重叠时,它们富集重叠相同的DMR,包括82个含有所有这四种因子的区域(图 3)c,d)。一种这样的DMR是在LRRC3,用全基因组显着性在WD肝特异性甲基化,并含有重叠为这些因素所有四个(图肝结合位点 3 E)。

WD特异性高甲基化肝脏DMR富含肝脏相关转录因子结合位点。一个 WD肝脏的DMR使用LOLA可用肝转录因子芯片起峰重叠从ENCODE和比值比作图并通过高甲基化DMR比值比来分类的。b使用HOMER测试WD肝脏DMR序列的富集的已知转录因子基序,并绘制具有肝脏ChIP-seq数据的因子。c与顶部肝脏转录因子ChIP-seq峰重叠的高甲基化WD肝脏DMR重叠。d转录因子重叠富集。e重组转录因子结合位点和高甲基化DMR的UCSC基因组浏览器视图LRRC3
WD进展相关的肝脏DMR显示补体激活和胚胎发育途径的失调与疾病严重性的增加
为了确定与WD进展相关的肝脏DNA甲基化变化,早期(0-1阶段)和晚期纤维化阶段(≥2)WD患者进行了比较并评估了DMR(早期WD n = 3,晚期WD n = 7) 。1339米高甲基化和低甲基化1119的区域被确定为与疾病进展变化的,受试者可以通过级甲基化这些的DMR的基础上进行分离(图 4的a,b)。然而,甲基化与协变量无关(附加文件2:表S6)。在这些WD进展DMR中,15个达到全基因组显着性(附加文件1:表S7)。这些重要DMR附近的基因包括参与血液凝固的基因(F7,F10),免疫激活(RNF123,SIGLEC15)和线粒体功能(KAT2A,LIAS和SARDH)。所有WD进展DMR的功能富集分析显示,高甲基化区域富集参与补体激活,体液免疫应答和脂质稳态的基因,而低甲基化区域富集参与心脏,呼吸,肌肉和内胚层发育的基因(图2)。 4 c)。这些数据表明在WD进展期间发生与补体激活和胚胎发育相关的基因的DNA甲基化改变。

肝脏DMRs区分早期和晚期WD患者。WD进展相关的肝脏DMR富集参与补体激活和胚胎发育的基因。高甲基化的DMR在晚期WD中具有较高的甲基化,而低甲基化的DMR在晚期WD中具有较低的甲基化(早期WD n = 3,晚期WD n = 7)。在早期与晚期WD肝脏DMR中使用甲基化的热图。绘制每个样品相对于每个DMR的平均甲基化的甲基化百分比。b在早期与晚期WD肝脏DMR中使用甲基化的主成分分析。省略号显示95%的置信区间。C与背景相比,早期与晚期WD肝脏DMR的GREAT功能富集分析的结果。显示了来自FDR q <0.05的高甲基化或低甲基化DMR的基因本体数据库的前10个术语(虚线表示显着性阈值)。对于这个以及随后的所有数据:FDR; 错误的发现率
WD相关的肝脏DMR基因富含药物靶标
鉴定与WD诊断和严重程度相关的肝脏中具有差异甲基化的基因提供了预测可能与这些基因相互作用并可能改变WD病理学的治疗剂的机会。为了鉴定与WD DMR相关的药物,已知药物 - 基因相互作用从药物基因相互作用数据库[ 18 ]获得并与WD DMR基因重叠(附加文件2:表S8)。最显着富集的药物是bepridil,它是一种钙拮抗剂,已知会影响线粒体功能[ 19 ],它与4种基因在早期WD中的差异甲基化相互作用:ATP1A1,CACNA1H,TNNC1和VIPR2(p = 1.2E-4)。WD特异性肝脏DMR基因显着富集与美法仑的相互作用,后者已被用于治疗肝脏转移[ 20 ],并与FANCC,GSTP1,MGMT,OPLAH和PLAT基因相关(p = 6.5E-4)。虽然这些药物对WD病理学的影响无法根据此分析进行预测,但这些数据确实提出了在WD模型系统中进一步研究的化合物。
WD相关的肝脏甲基化不受细胞类型变化的影响
在诸如肝脏的大块组织中观察到的DNA甲基化变化可受到样品之间各个细胞类型的比例差异的影响。为了确定细胞类型的变化是否影响本研究中鉴定的DMR,我们检测了人肝细胞(HEP),肝星状细胞(HSC)或肝窦上皮细胞(LSEC)中特异性高甲基化的启动子组[ 21 ]。在所有已鉴定的WD DMR中,只有63种WD特异性,3种早期WD特异性和86种WD进展DMR与细胞类型特异性甲基化启动子重叠(附加文件1):图S6a)。来自每次比较的少于5%的DMR位于细胞类型特异性甲基化启动子。在用于鉴定DMR的所有样品组之间比较细胞类型特异性启动子的甲基化百分比(附加文件1:图S6b-d,附加文件2:表S9)。大多数细胞类型特异性启动子在任何样品组之间没有差异甲基化。与HC相比,WD样品在7%的HEP启动子,4%的HSC启动子和7%的LSEC启动子上名义上差异甲基化(p <0.05)。与早期WD相比,在晚期观察到最大的影响,其中8%的HEP启动子,9%的HSC启动子和13%的LSEC启动子名义上差异甲基化(p <0.05)。这些结果表明,细胞类型比例的改变对WD肝脏中观察到的差异甲基化没有主要影响。
血液中的DMR将WD与对照区分开来
虽然肝脏WD特异性DMR的鉴定与疾病见解最相关,但我们询问是否可以在更易获得的血液样本中鉴定出类似的表观基因组特征作为潜在的WD生物标志物。在两个独立的群组中共有82个全血样品可用于DNA提取和甲基化组分析(附加文件1:表S10,附加文件2:表S11)。WD患者均在诊断时招募,而不是抗铜治疗。对于NAFLD和PSC受试者,从无纤维化到肝硬化,可直接从肝脏活组织检查或非侵入性评估获得纤维化阶段。WD的全血甲基化组(n = 40),HC(n = 12),并且通过WGBS评估DC(其包括NAFLD和PSC患者,n = 20),并且针对每个成对比较调用DMR(图 5a)。鉴定出187个高甲基化和75个低甲基化的WD特异性DMR,区分WD与HC和DC患者。在这些WD-特定血液的DMR使用甲基化水平,大部分患者从WD HC和DC患者分离(图 5 B,C)。相反,大多数DMR中的甲基化与人口统计学协变量无关(附加文件2:表S12)。

血液DMR区分WD患者与对照组。鉴定了 WGBS衍生的DMR,其区分WD与HC和DC,但不区分来自HC的DC(WD n = 40,HC n = 12,DC n = 20)。b使用WD特异性血液DMR中的甲基化,HC,WD,NAFLD和PSC血液的热图。绘制每个样品相对于每个DMR的平均甲基化的甲基化百分比。c在WD特异性血液DMR中使用甲基化的主成分分析。省略号显示95%的置信区间
由于肝脏增强子中肝脏WD特异性DMR的富集,还评估了血液WD特异性DMR中的染色质状态富集(附加文件1:图S7)。总体而言,DMR富集了免疫细胞增强子区域,尤其是在造血干细胞中预测的那些,其重叠51%的DMR(优势比= 9.1,FDR q = 6.1E-58)。另外,高甲基化区域在单核细胞,中性粒细胞,B细胞和自然杀伤细胞增强剂中富集更多,而低甲基化区域在T细胞增强剂和启动子中更富集。
血液中的DNA甲基化反映了WD特异性肝脏DMR的一个子集
正如预期的那样,血液中鉴定的WD特异性DMR的数量远低于肝脏中鉴定的数量。然而,血液DMR附近的99个(22%)基因与肝脏中鉴定的基因重叠,显着富集(Hyper:p = 1.1E-25,Hypo:p = 2.0E-6,附加文件1:图S8a) 。在血液和肝脏中具有重叠位置和共同方向的10个DMR附近的基因包括CSAD,ITGB7,LSM12,PSMD9,RRN3P2,SNX29P2和PFN3。特别是,HDAC5内含子和LSM12上游的肝脏DMR在WD肝脏和血液中特异性地和显着地高度甲基化(附加文件1:图S8b-d)。
人和小鼠WD DMR基因在组织内和组织之间重叠
为了确定与ATP7B功能丧失相关的DMR基因是否在人和小鼠之间保守,肝脏和血液中的人DMR基因与先前在WD模型tx-j小鼠的胎肝中鉴定的DMR基因重叠,与野生型C3H小鼠相比较。 (附加文件1:图S9,附加文件2:表S13)[ 15 ]。在人肝脏和小鼠胎肝中,20个基因在相同方向上差异甲基化(hyper:q = 7.7E-2,hypo:q = 4.7E-4)。在ATP7B功能丧失时具有保守差异甲基化的基因中有ALKBH5,GRID2IP,KDELR2,CACNA1H,CAMK2B和WNT11。在血液中,7个基因在与小鼠胎肝相同的方向上差异甲基化(hyper:q = 6.6E-3,hypo:q = 1.3E-1)。其中,ZNF750和MAD1L1在人肝脏中也是差异甲基化的。这些基因的保守差异甲基化表明它们代表了WD病理学中重要的早期扰动。
血液中的DMR区分患有肝脏和神经系统WD表型的患者
表型变异是WD的一个重要特征,不能通过基因型解释,因此我们分析了血液中的甲基化,以确定是否存在分化WD肝病(WDH)和神经系统(WDN)症状的WD患者(WDH n = 20,WDN n) = 20)。总共1346个区域的甲基化了,而1514个区域被在WD患者肝症状低甲基化相比,那些与神经症状(图 6的a,b)。患者可以通过表型清楚地聚集,表明血液中存在与WD症状相关的表观遗传差异。相反,WD表型DMR中的甲基化与协变量无关(附加文件2:表S14)。所有DMR附近的基因都富集了异常B细胞生理学中的功能,而WDH中低甲基化区域附近的基因富含痴呆,精神衰退和属于染色质修饰多梳抑制复合物1(PRC1)的功能(图6)。 C)。类似于WD特异性血液DMR,WD表型DMR富含免疫细胞增强剂,尤其是造血干细胞中的那些(附加文件1:图S7c,d)。独特地,WD表型DMR富含二价增强子,其由H3K4me1和H3K27me3定义并由PRC1调节,并且特别富集在T细胞中(优势比= 4.6,FDR q = 1.5E-171)。

)
血液DMR区分患有肝(H)或神经(N)WD的患者。鉴定了区分WDH和WDN的DMR。一个使用甲基化WDH与WDN血液的DMR热图。绘制每个样品相对于每个DMR的平均甲基化的甲基化百分比(WDH n = 20,WDN n = 20)。b使用WDH中甲基化与WDN血液DMR的主成分分析。省略号显示95%的置信区间。c与背景相比,WDH与WDN血液DMR的GREAT功能富集分析的结果。来自FDR的基因本体数据库的术语q 显示<0.05,虚线表示显着性阈值。“所有DMR”(绿色)指高甲基化和低甲基化DMR; “低甲基化DMR”(蓝色)是指与WDN相比在WDH中具有较低甲基化的DMR
WD表型血液DMR在独立队列中对肝脏和神经系统WD进行分类
使用基于血液的生物标志物区分WD表型在患者诊断中具有潜在的应用。为了评估所鉴定的WD表型血液DMR的准确性以将WD患者分类为肝脏或神经形式,在原始患者群组中对Adaboost分类器在这些区域中的血液甲基化进行训练并且在独立群组上进行测试(WDH n = 5) ,WDN n = 5)。识别出44个DMR,其特征重要性大于零,并用于模型中(图 7a,b)。这些DMR附近的基因包括几个具有已知神经功能的基因,如CHRNB3,CYP46A1,GALR1,LHX4,NGF,NPTXR,SHH和WNT7B,和肝功能,包括ONECUT1(图 7 C,附加文件2:表S15)。重要的是,分类器能够正确地识别除了一个独立测试样品之外的所有样品,其具有WD的肝脏或神经形式(精确度= 0.9,曲线下的接收器操作特征(ROC)面积(AUC)= 0.8,图3)。 7 d)。这些结果表明,在这44个区域中使用血液DNA甲基化的患者中可以鉴定出WD表型。
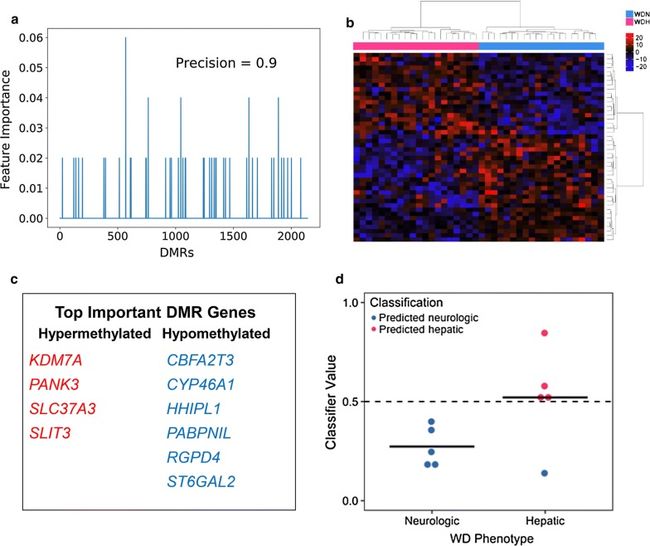
选择的WD表型血液DMR在独立的测试组中对来自神经系统WD的肝脏进行分类。使用AdaBoost算法的分类器用WD患者血液中WD表型DMR的甲基化值训练(WDH n = 20,WDN n = 20)。a训练后2860个DMR中的每一个都确定了特征重要性。b在训练样本中使用甲基化的分类器中使用的44个DMR的热图。c特征重要性≥0.04的DMR附近的基因。d分类结果为WD患者的测试队列。在图中,该列代表真实的表型,而颜色代表预测的表型(WDH n = 5,WDN n = 5,精度= 0.9,ROC AUC = 0.8)
讨论
这是第一项通过WGBS在全基因组水平上研究WD患者甲基化变化的研究。这种表观基因组特征揭示了肝脏特异性增强子和转录因子结合位点发生的甲基化变化,表明WD是一种遗传性疾病,其进展和潜在的发病机制与表观遗传变化有关。与健康受试者和患有其他肝病的受试者相比,我们在WD患者中证实了特定的肝脏发病相关DNA甲基化标记。在来自不同队列的患者的肝脏和血液中鉴定了这些DNA甲基化差异的子集,支持血液中的甲基化标记物可以反映另一组织中的疾病发病机理的前提。此外,DNA甲基化差异也基于肝脏与神经系统表现区分患者,特别是编码具有机械意义的表观遗传因子的基因,例如PRC1中的那些。这些区域的一小部分可以在独立的队列中对WD患者表型进行分类,证明了DNA甲基化的诊断潜力。WD的这种表观基因组特征揭示了对基因途径,基因 - 药物相互作用和潜在生物标志物的见解,这些可能在临床上用于WD的早期干预和治疗。证明了DNA甲基化的诊断潜力。WD的这种表观基因组特征揭示了对基因途径,基因 - 药物相互作用和潜在生物标志物的见解,这些可能在临床上用于WD的早期干预和治疗。证明了DNA甲基化的诊断潜力。WD的这种表观基因组特征揭示了对基因途径,基因 - 药物相互作用和潜在生物标志物的见解,这些可能在临床上用于WD的早期干预和治疗。
我们之前在WD的tx-j小鼠模型中的数据显示,胎儿肝脏中肝细胞生长和成熟缺乏的全面减少,通过母体膳食补充甲基供体胆碱来减轻[ 22 ]。在tx-j小鼠的胎肝中也观察到差异甲基化,其通过补充胆碱改善[ 15 ]。在患者肝脏中鉴定的差异甲基化基因与tx-j胎儿肝脏中的差异甲基化基因显着重叠,并且包括编码在肝脏发育中重要的转录因子的几个基因。GATA6参与肝脏成熟的多个阶段,并在胎儿早期发育过程中在肝细胞中高表达[ 23 ]。FOXA1在肝脏胚胎发生过程中启动规范是至关重要的[ 24 ]并且也在成年肝脏中表达,而VTN,一种FOXA1靶基因,参与胚胎干细胞成熟[ 25 ]并且已经显示在胎儿肝星状细胞中被上调[ 26 ]。GATA6,FOXA1和的Vtn还发现在成年TX-J肝脏差异表达。WD的这些DNA甲基化特征指向肝脏发育和再生途径,这些途径又可能受胎儿和成年生命中甲基供体饮食因素的影响。
在WD中鉴定的高甲基化DMR特异性地富集在肝特异性增强子和转录因子结合位点中,这是WD领域的新发现。肝特异性增强子中的高甲基化DMR可反映增强子使用或HNF4A结合的特定缺陷。HNF4A是肝细胞成熟的关键转录因子,是维持肝细胞上皮表型所必需的[ 27 ]。之前的一份报告显示,WD患者HNF4A和RXR与启动子反应元件的结合受损[ 28 ]。此外,肝脏X受体/类视黄醇X受体的失调与WD的发病机制有关 [29]]。发现HNF4A,RXR,FOXA1和FOXA2的肝结合位点在LRRC3的启动子中显着高度甲基化的WD特异性肝DMR处重叠。与早期WD相比,LRRC3的另一个DMR在晚期显着高度甲基化。以前,LRRC3与铂诱导的肝损伤和氧化应激的易感性有关[ 30 ]。我们的结果表明HNF4A和肝脏特异性增强子和启动子中的其他转录因子结合位点的高甲基化为WD发病机制中的这些先前分子发现提供了新的潜在解释。
另外,从WD的这个表观研究得到的见解可对有争议铜累积和潜在的降低肝癌风险之间在WD [关系光棚31,32 ]。最引人注目的发现之一是肝脏和血液之间重叠的DMR基因在癌症的表观遗传机制中起主要作用。特别是,HDAC5活化与肝细胞增殖和肝癌发生有关[ 33 ],并参与脂质代谢和脂肪酸氧化[ 34 ]。无论NCOR2和CTBP2在WD患者的肝脏和血液中甲基化和功能在癌症失调转录共抑制。NCOR2与两个核受体,包括LXR和RXR,和IIa类HDAC的,如HDAC5 [相互作用35,36 ]。NCOR2在膀胱癌,乳腺癌和前列腺癌中也有差异表达[ 35 ]。CTBP2的功能是在分化期间募集PRC2蛋白以添加H3K27me3,并且还与HDAC相互作用[ 37 ]。此外,CTBP2在前列腺癌中过表达并与肿瘤进展相关[ 38 ]。其他基因,包括GATA2,GADD45B和MIR126,WD肝脏和血液中的差异甲基化,也在癌症的表观遗传机制中发挥作用。WD中这些表观遗传调节因子的失调可能会影响这些患者对癌症的易感性。
先前的几项研究试图将全血或外周血单核细胞(PBMC)中特定的DNA甲基化变化确定为肝病的指标。特别是,对NASH患者的分析发现了与胶原蛋白含量相关的基因[ 11 ]和脂肪变性[ 39 ]的DNA甲基化变化]。我们的深入分析确定了可在诊断时区分早期和晚期WD和肝脏与神经系统表型的DMR。据我们所知,这是第一项确定发病机制 - 有意义标志物组合的研究,以及与临床和遗传参数相关的诊断潜力。在我们对早期WD肝脏样本的晚期分析中,我们发现了补体途径中涉及的高甲基化基因,这些基因以前被证明是WD的血清生物标志物 [40]。]。我们的研究描述了一组来自全血的差异甲基化基因与WD肝脏中鉴定的基因重叠,表明这些基因座可以作为WD在未来研究中疾病进展的各个阶段的潜在血液生物标志物。
该研究的局限性包括使用全血和全肝组织,因为两者都是许多不同细胞类型的混合物。在肝脏中评估细胞类型的影响,其中发现在特定肝细胞类型中大多数高甲基化的启动子与WD肝脏中的差异甲基化无关。第二个限制是健康体重的个体缺乏肝脏控制。通过检查与DMR甲基化无关的BMI来控制这一点。与脂肪酸氧化和脂质分解代谢相关的途径中基因的DNA甲基化的变化与WD动物模型中下调的脂质代谢途径的先前证据一致[ 8]。第三个限制是甲基化分析的覆盖率相对较低,这可能限制了检测所有可能的甲基化差异的能力。然而,相对大量的样本允许我们捕获大量的DMR,这些DMR在独立群组中得到验证,提供了WD中DNA甲基化作用原理的证据。此外,所述测序方法的显着成本可被视为DMR标记物作为WD诊断测试的未来适用性的限制因素。使用特定WD预测性DMR的减少表示将显着降低成本,增加测序覆盖率,并使其成为临床实践的可访问技术。
结论
WD代表一种非常独特的遗传条件,其中ATP7B中的突变与铜,饮食和代谢相互作用,影响其复杂和变化的表型,因此血液或其他可及组织中疾病进展的表观遗传指示显然在临床上是有用的。这些DMR可能代表WD的表观基因组特征,包括铜水平升高,疾病进展反应和饮食等环境输入的直接改变。此外,对编码药物可靶向表观遗传修饰因子(如HDAC5)的基因的DNA甲基化变化的鉴定可揭示对WD治疗中现有药物的再利用的见解。
方法
人体肝脏活组织检查
来自WD患者的样品获自Heidelberg大学(德国海德堡)和加州太平洋医学中心Ibrahim El-Hefni Biorepository(美国加利福尼亚州旧金山)。来自海德堡大学的肝脏样本来自接受肝脏移植的WD患者的外植肝脏。来自加利福尼亚太平洋医疗中心的肝脏样本是通过经皮肝脏活组织检查获得的,用于诊断和分期目的。来自DC和HC受试者的肝样品获自加利福尼亚太平洋医疗中心和加州大学戴维斯GI生物库(附加文件1:表S1,附加文件2):表S2)。DC患者的经皮肝脏活检取自NAFLD。在患有非糖尿病的受试者的减肥手术时获得HC肝脏活组织检查,脂肪变性小于5%且组织学上无炎症。如前所述,活组织检查对炎症,脂肪变性和炎症进行分级[ 41 ]。
人体全血样本
来自患有WD的患者(与提供肝脏活组织检查的患者不同的组)的样品获自精神病学和神经病学研究所(波兰华沙)。HC样本来自华沙和萨克拉门托当地社区的志愿者。DC样本来自诊断为NAFLD和PSC的患者,他们在加州大学戴维斯分校肝病学诊所连续出现进行评估,并同意提供全血用于DNA提取,以存储在加州大学戴维斯分校生物库中。选择NAFLD和PSC作为DC,因为在WD的鉴别诊断中应考虑这两种条件,NAFLD患者呈现表观遗传变化,并且PSC也与铜积累相关。在HC和WD组中,全血样本按年龄,性别和BMI进行匹配(附加文件1:表S10,附加文件2:表S11)。患有其他慢性肝病(包括乙型肝炎或丙型肝炎,自身免疫性肝炎,酒精性肝病,原发性胆汁性胆管炎,血色素沉着症)的患者被排除在外。根据莱比锡标准确定WD患者在诊断时被招募,因此不进行抗铜治疗。选择测试组中的WD患者具有ATP7B H1069Q错义突变。根据患者的流行表型将患者分类为肝脏或神经系统表现。将所有样品去标识,在干冰上运输,并储存在-80℃下用于进一步分析。
全基因组亚硫酸氢盐测序(WGBS)和分析
使用TruSeq DNA甲基化试剂盒(Illumina,San Diego,CA,USA)从亚硫酸氢盐转化的DNA制备WGBS文库。来自肝脏和血液训练样品的文库在HiSeq4000上测序,而来自血液测试样品的文库在HiSeq2000(Illumina,San Diego,CA,USA)上测序。将读数与hg38参考基因组比对,并在HC,WD和DC样品之间鉴定DMR。WGBS数据分析的所有代码都可以在GitHub上获得(https://github.com/cemordaunt/WilsonDiseaseEpigenome)。
关于WGBS数据分析和小鼠模型研究的其他方法在附加文件1:补充方法中给出。
非酒精性脂肪性肝病(NAFLD)和原发性硬化性胆管炎(PSC)