精神分裂型患者大脑结构和功能连接的改变
背景:精神分裂型指的是在一般人群中低于临床阈值的精神分裂样特征。精神分裂症的病理发展被假设为从最初的脑连接断开和脑连接补偿共存到脑连接失代偿的演变过程。
方法:在本研究中,我们结合脑白质结构连通性、弥散张量成像数据的静态和动态功能连通性分析和静息态功能磁共振成像数据,研究了与分裂型相关的脑连通性变化。共有87名分裂型性状高水平的被试和122名对照组完成了实验。研究了264个脑兴趣子区域在全脑白质结构连接概率、静态平均功能连接强度、动态功能连接变异性和稳定性方面的组间差异。
结果:我们发现高分裂型个体在任务控制网络内和默认模式网络内的结构连通性概率增加;默认模式网络内和听觉网络与皮层下网络之间功能连接的变异性增加,稳定性降低;静态平均功能连接强度的下降主要与感觉运动网络、默认模式网络和任务控制网络有关。
结论:这些发现突出了与分裂型相关的大脑连通性的具体变化,表明在全脑功能断开的情况下,默认模式网络和任务控制网络的结构连通性的失代偿性和代偿性变化可能是高度分裂型个体的一个重要的神经生物学相关性。
1.简介
精神分裂类型指的是在一般人群中发现的亚临床精神分裂样特征。其临床表现包括认知知觉(阳性)、人际关系(阴性)和认知紊乱特征,这些症状在质量上与精神分裂症症状相似,并与精神分裂症责任相关。分裂型可能代表精神分裂路径上的一种内表型,可以使用分裂型人格问卷调查。流行病学研究表明,在一般人群中具有高水平分裂型特征的个体比例约为10%。研究分裂型有助于理解精神分裂症相关的精神病理学和推定的精神分裂症内表型。
此前的研究表明,精神分裂症的神经生物学基础可能是大脑连接断开或大脑连接障碍。具体来说,一项研究发现,在一般人群中,额颞白质束的分数各向异性值与SPQ认知知觉亚量表的得分呈负相关。静息状态功能磁共振成像(rs-fMRI)研究发现,分裂型特征水平高的个体楔前叶和海马旁区之间的功能连通性降低,与SPQ总分呈负相关。在一般人群中,SPQ认知知觉亚量表得分与腹侧纹状体与额叶皮层之间的功能连接呈正相关,与背侧纹状体与后扣带皮层之间的功能连接呈负相关。一项独立成分分析研究发现,在一般人群中,SPQ认知知觉亚量表得分与视觉网络的低频振荡呈正相关,而SPQ人际关系亚量表得分与听觉网络的低频振荡呈负相关。这些发现表明分裂型的发展可能与大脑网络连接的改变有关。对具有高水平分裂型特征的个体进行全面调查,可以加深我们对与分裂型相关的结构和功能连通性变化的异同的理解,并可以确定哪些变化在高度分裂型的个体中是重要的。
以往的分裂型研究也很少关注功能连通性的时间动态。大脑子区域之间的时间耦合是时变的,而不是时不变的。动态功能连接(Dynamic functional connectivity, DFC),也被称为连接体(chronnectome),可以深入了解交互功能连接网络的可变性。此前的一项研究发现,精神分裂症患者在丘脑高连通性和感觉低连通性方面表现出动态变化。另一项研究发现,有精神类经历的个体表现出与视觉网络的超连通性和默认模式网络的低连通性相关的大脑动态变化。总的来说,高分裂型特征个体的DFC稳定性值得进一步研究。
在本研究中,我们利用弥散张量成像(DTI)和静息态功能磁共振成像(rs-fMRI)技术研究了结构连通性、静态功能连通性和DFC。我们通过概率追踪成像和新的可靠的动态条件相关(DCC)方法,该方法比滑窗和非参数估计方法更可靠,对时变特征进行了定量描述。我们在本研究中使用的代表性全脑模板由Power等人(2011)确定,其中包括属于不同大脑网络的264个感兴趣区域(ROI)。计算了各roi之间的结构连接概率、静态平均功能连接强度和DFC指标(包括DFC变异性和DFC稳定性)。
基于先前关于与精神分裂症谱系障碍发展相关的大脑区域或网络之间的缺陷和代偿性大脑连接变化的发现提出了一种精神分裂症神经生物学发展可能性,称为连通性失代偿假说,该假说假设精神分裂症谱系连续体的病理发展是一个从功能代偿到部分失代偿再到完全失代偿的渐进过程。在该模型中,由于遗传和/或不良内外环境因素的综合作用,易感个体中与情绪、感觉和知觉推理相关的一些神经回路可能会逐渐受损。这些异常仍然可以在最初得到部分补偿,这些个体可以通过逐渐的结构增殖、再植、适应性功能调整或与认知调节和自我监控相关的大脑子区域重组来维持正常功能。他们中的大多数人永远不会得这种病。然而,如果这些诱发因素过度且持续存在,或者神经补偿机制失效,可能会进一步加重精神类特征和认知功能失调,导致临床发病。具有高水平分裂型特征的个体处于这一过程的早期阶段,对他们的调查可以提供早期干预和预防的最佳机会,并避免混杂因素,如抗精神病药物暴露和精神病共病。
因此,我们的研究目标是:(1)与对照组相比,研究分裂型特征高水平个体在功能和结构连通性方面的共同和特定变化;(2)在分裂型高水平个体中验证连通性失代偿假说。如果高分裂型组roi之间的功能和结构连通性均增加,则可认为是该通路中的一种补偿效应。如果roi之间的功能连通性降低而结构连通性增加,可能意味着失代偿。基于之前的研究,我们假设高分裂型特征个体的结构连接和功能连接的变化会同时表现出失代偿和代偿效应,这可能预示着大脑连接的断开。
2.方法和材料
2.1 被试
2018年,我们首先通过广告方式从北京各高校随机收集了4000份在线分裂型人格问卷(SPQ)。每位志愿者都是一名大学生,都有报酬。将这些参与者SPQ总分的第90百分位(41分)作为分值,这与之前的研究一致,将一般人群中SPQ得分前10%的人作为高分裂型组。分数在41分以上的学生被分到高分裂型组,分数在41分以下的学生被分到对照组。然后,被分配到高分裂型组的个体和相应数量的对照组通过短信联系,邀请他们参加后续的MRI实验。其中90%的参与者对在线调查做出了回应。其中246名应答者同意进行脑部扫描。所有受试者的入选标准均为18岁及以上的汉族。所有参与者的排除标准包括任何身体或精神障碍和物质依赖的诊断,任何精神障碍的家族史和任何MRI扫描禁忌症。此外,没有完成所有研究步骤、成像质量较差、扫描时头部运动大于2mm和/或2°的受试者被排除在外。根据这些标准,35名参与者被排除在外。该研究获得了中国科学院大学心理学系伦理委员会的批准。所有参与者均给予书面知情同意。
2.2 问卷
SPQ是一个74个项目的自我评估量表,用于评估分裂型特征。这些项目可以聚为三个因素,即认知-知觉因素、人际因素和无序因素。中文版SPQ具有良好的心理测量特性。所有参与者的智商(IQ)采用中文版韦氏成人智力量表修订本的简写形式进行估计。参与者被要求在MRI扫描前再次填写SPQ,这样他们的SPQ分数就会在智商评估的同一天被确定。
2.3 MRI采集
在参与者完成问卷评估后,使用中国科学院心理研究所2018 - 2019年的3.0 T GE MR扫描仪依次获取静息态功能磁共振(rs-fMRI)、解剖和弥散张量图像(DTI)。
2.4 DTI预处理
DTI数据用FSL (https://fsl.fmrib.ox.ac)和PANDA 1.3.1 (http://www.nitrc.org/ projects/)进行预处理。处理过程与我们的另一项精神分裂症DTI研究相同。这些数据首先由一位有经验的放射科医生进行筛选,以排除偶然的结构异常。两名参与者被排除在外。然后使用FSL的fslroi命令裁剪原始图像。然后对扫描过程中涡流引起的DWI畸变和头部运动进行校正。通过仿射变换将DWI注册到b0图像。个体空间图像标准化到MNI空间的FMRIB58_FA模板,6mm高斯核平滑。然后使用test bvecs实用程序找到正确的倒置和交换参数的DWI方向。使用BedpostX建立每个体素上扩散参数的分布,并创建运行下一个概率束造影程序所需的所有文件。采用以下参数在半径为6 mm的264个roi之间进行概率束成像:曲率阈值= 0.2,最大步数= 1000,步长= 0.5 mm,样本数= 50 000,路径分布的跟踪类型。对于每个ROI,通过从特定ROI中的所有体素中播种来执行概率束成像。从ROI i到ROI j的白质连接概率定义为通过ROI的纤维数量除以从ROI采样的纤维总数。为每个参与者生成一个264 roi概率矩阵。然后采用0.001的阈值来减少roi之间的假阳性连接。由于区域i到区域j的连接概率不一定等于区域j到区域i的连接概率,因此对这两个概率取平均值,得到每个参与者对称的无方向连接矩阵,得到连接概率值34 716个。
2.5 Rs-fMRI预处理
Rs-fMRI数据预处理采用统计参数映射软件(SPM12, https://www.fil.ion.ucl.ac.uk/spm/)、脑成像数据处理与分析(dabi4.0, http://www.rfmri.org/dpabi)和脑小波1.1 (http://www)。软件在MATLAB R2016b (MathWorks Inc., https://www.mathworks.com)运行。前10卷被丢弃。然后分别用dabi和脑小波进行切片定时、再排列和小波去噪。随后,回归出包括Friston 24头部运动参数、白质信号和脑脊液信号在内的讨厌协变量。结构图像与前、后连合线人工共配并对齐,与平均功能图像共配,并在SPM中使用DARTEL分割为灰质、白质和脑脊液。然后将每个功能图像归一化到蒙特利尔神经学研究所标准空间的333mm³体素中,然后对图像进行带通滤波(0.01-0.1 Hz)。以6 mm半高宽核进行空间平滑。平均帧位移(FD)作为协变量被纳入后续分析。提取264个roi的时间序列,每个参与者的半径为6 mm。使用dpabi 4.0中的Pearson相关构建静态fMRI网络,使用MATLAB 2016b中Martin Lindquist动态连接工具箱中的动态条件相关(DCC)脚本构建动态大脑网络(https://github.com/canlab/ Lindquist_Dynamic_Correlation)。生成每个参与者的264 ROIs - 264 ROIs静态功能连通性矩阵和264 ROIs - 264 ROIs - 290时间序列DFC矩阵。计算两个roi之间每个连接的平均功能连通性强度、DFC标准差和DFC稳定性,得到34 716个值。DFC标准差(变异性)表示DFC随时间的总体波动水平。DFC稳定性描述了相邻时间序列之间DFC的时间变化的连续性。
2.6 数据分析
采用基于网络的统计软件和Matlab 2016b,对两组间的平均功能连接强度、DFC变异性、DFC稳定性、白质连接概率进行ANCOVA分析,并在错误发现率(FDR)为0.05的基础上,回归性别、年龄、智商(FD值也作为功能连接分析的协变量)。结果图像用BrainNet Viewer可视化。最后,使用matlab 2016b中的fdr_bh函数,对具有显著组间差异的连接概率和功能连通性值与高分裂型组的SPQ评分进行Pearson相关分析,同时回归性别、年龄、智商(以及功能连通性分析中的FD),显著性水平设置为fdr 0.05校正。
- 结果
经过筛选和质量控制,最终纳入87例高分裂型个体和122例对照组。所有受试者的平均年龄为21.44岁,IQ和FD分别为124.89,0.05。两组在性别、年龄、智商和平均FD方面差异无统计学意义。两组在SPQ得分上差异显著。
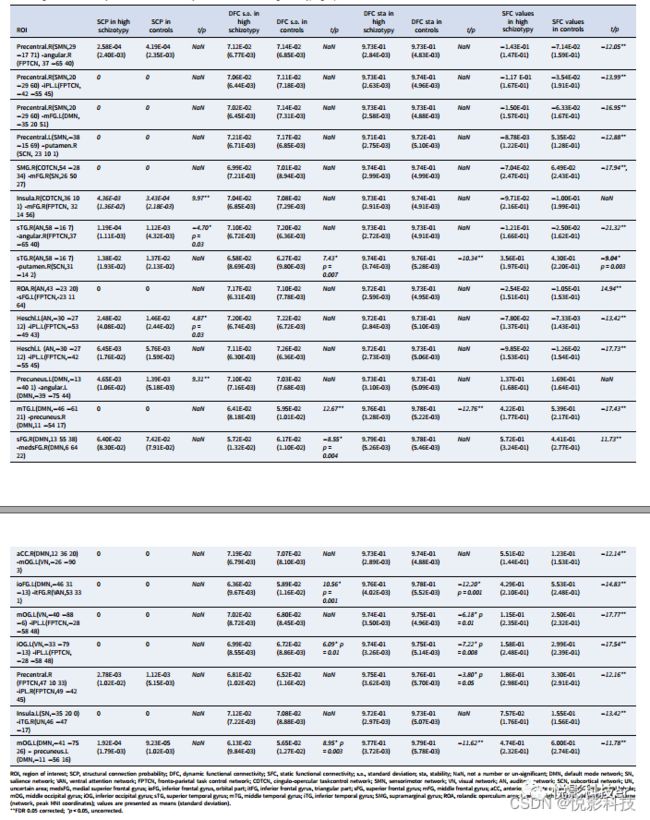
与对照组相比,高分裂型组在右侧脑岛(扣带-顶盖任务控制网络,COTCN)和右侧额中回(额顶任务控制网络,FPTCN)之间的白质连接概率增加;左楔前叶之间(默认模式网络;DMN)和左角回(DMN);右侧楔前叶(DMN)和左侧颞中回(DMN)之间的DFC变异性增加,稳定性降低;以及右侧颞上回(听觉网络)和右侧壳核(皮层下网络)之间以及左侧楔前叶(DMN)和左侧枕中回(DMN)之间的稳定性下降。
此外,18个平均功能连通性强度值在组间存在显著差异。高分裂型组与对照组相比减少了16个,主要涉及感觉运动网络、听觉网络、视觉网络、DMN和任务控制网络。在左侧额上回(FPTCN)和右侧圆状盖区(听觉网络)之间,以及在右侧额上回(DMN)和右侧内侧额上回(medsFG, DMN;见表1和图1)。
高分裂型组的相关分析发现,SPQ认知知觉亚量表得分与左侧枕中回(视觉网络)与左侧顶叶下小叶(FPTCN)的平均功能连接强度呈负相关。
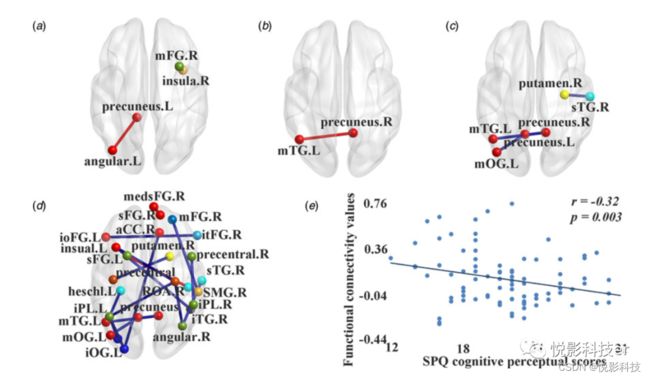
图1 高分裂型个体与对照组之间的白质连接概率和功能连接值有显著差异。
表1 高分裂型组与对照组之间存在显著的白质和动态功能连通性差异
4.讨论
在这项研究中,我们调查了具有高水平分裂型特征的个体的全脑结构和功能连通性的变化。我们发现在高分裂型组中,DFC变异性增加,DFC稳定性降低,后DMN子区域和任务控制网络内部的静态功能连接强度降低,听觉网络和任务控制网络之间的静态功能连接强度降低。同时,在高分裂型组中,两者之间的结构连通性显著增加。这些结果支持了我们在高分裂型个体中的连通性失代偿假设。然而,我们也发现,在高分裂型组中,DMN前部子区域内、FPTCN与听觉网络之间的静态功能连接强度增加,FPTCN与听觉网络之间的结构连接增强,这可能是代偿效应。同时,我们发现颞上回(听觉网络)和角回(FPTCN)之间的静态功能连接和结构连接都减少了。它可能代表神经通路受损。最后,我们没有发现任何既增加功能连通性又减少结构连通性的通路。我们的发现可以扩展我们对大脑子区域之间的时间动态耦合的理解,并可能为深入了解高分裂型组。此外,我们的研究结果支持我们的假设,即在分裂型高水平个体中,大脑连接失代偿和大脑连接补偿共存。
先前的元分析研究报道精神分裂症患者主要表现为结构连通性的减少,特别是在额叶和颞叶。相反,我们发现与对照组相比,高分裂型的个体在右侧脑岛(COTCN)和右侧额叶中回(mFG, FPTCN)之间以及左侧楔前叶(DMN)和左侧角回(DMN)之间表现出更高的白质连接概率。角脑回和脑岛整合并连接周围的许多区域,这些区域之间结构连接的增加可能是一种补偿与突触可塑性和轴突增殖相关的机制,可缓解执行控制功能障碍和自我感觉整合。这些白质通路的代偿性改变是否有效,可以从它们的功能连通性变化来推测。
然而,我们也发现,增加的DFC变异性降低了DFC的稳定性,以及后DMN子区域和任务控制网络内的静态功能连通性强度,并且在高分裂型组中,SPQ认知知觉亚量表得分与视觉网络子区域和任务控制网络之间的平均功能连通性强度呈负相关。这些功能障碍可能与不稳定的大脑活动、紊乱的自我参照思维和意识障碍有关,可能是分裂型特征高水平个体的异常感知和怀疑的重要神经功能基础。再加上高分裂型组中它们之间的结构连接增加的结果,这可能表明在高分裂型特征的个体中,这些区域之间的大脑连接不一致,神经补偿无效,并支持精神分裂症谱系障碍发展过程中的后DMN内、听觉网络、视觉网络和任务控制网络之间的连接失代偿的存在。
此外,高分裂型组也表现出听觉网络、感觉运动网络和壳核子区域之间的DFC稳定性和静态功能连接强度下降;与对照组相比,主要是感觉运动网络、听觉网络、视觉网络、DMN与任务控制网络之间的静态功能连接强度下降。前者可能反映了处理传入感觉信号的无效多巴胺能传递在高分裂型个体中已经出现。后者可能表明听觉、视觉、知觉处理和认知控制能力之间的功能失调分离、自我意识和外部环境信息整合以及感知-行为整合的缺陷。这些变化可能是与包括认知障碍在内的早期精神病倾向相关的关键特征,以及偏好先验主观知识而非传入感官信息的倾向。此外,这些功能连通性特征降低的子区域大部分没有表现出任何结构连通性变化,这表明这些功能障碍可能是由灰质神经元的树突变化引起的,或由其他连通性介导的。
此外,我们发现DMN前部的子区域内,主要是额上回和medsFG之间,以及FPTCN子区域和听觉网络之间的静态功能连接强度增加,并且在高分裂型组中,FPTCN和听觉网络之间的结构连接增强。这些子区域之间连接的增加可能表明它们在高级认知调节方面的自适应努力,以缓解感觉运动和社会情绪功能障碍。这可能是维持高分裂型个体正常社会行为的重要神经生物学代偿机制。这些变化的神经化学基础可能是由于功能重叠的神经递质如多巴胺、谷氨酰胺、乙酰胆碱和5-羟色胺的增加,以及突触可塑性和轴突增殖。我们还发现在高分裂型组中,颞上回(听觉网络)和角回(FPTCN)之间的静态功能连通性和结构连通性都降低了。它可能表明与异常的听觉感觉监测和自我参照处理相关的轴突功能缺陷。
综上所述,这些结构和功能连通性的结果可能意味着DMN和任务控制网络内的连通性增强不能完全补偿分裂型特征高水平个体的感觉-知觉功能失调变化。n-甲基-d-天冬氨酸受体(NMDAR)功能减退而不是轴突病理引起的突触整合异常可能是高分裂型个体中大多数功能连通性下降的关键神经化学基础,这些变化可能与精神分裂症遗传风险变异有关,包括DISC1, PPP1R1B, PRODH和ZNF804A。同时,在精神分裂症患者中发现的白质结构改变可能是精神分裂症症状恶化的病理生理后果,而不是疾病的原因。
综上所述,我们的研究结果表明,高分裂型个体既表现出代偿性连通性,也表现出缺陷性连通性,主要与DMN、任务控制网络和感觉运动网络有关。我们的研究结果表明高分裂型个体的大脑连通性适应性变化,并为精神分裂症谱系障碍的连通性失代偿假说提供了可能的神经生物学基础。
参考文献:Altered brain structural and functional connectivity in schizotypy