集体心智:社会网络拓扑塑造集体认知
来源:集智俱乐部
作者:Ida Momenneja
译者:许逸聪
审校:陈斯信
编辑:邓一雪
导语
人类是社会动物,人类的认知也是通过在社会网络中学习和记忆而形成。人类的社会网络与其他物种不同,有着复杂多样的成分和拓扑,这样的社会环境塑造了人类特有的集体认知。这篇综述回顾了支持“社会网络拓扑塑造集体认知及行为”这一假说的相关研究。文章探讨了社会网络拓扑如何影响人类集体记忆、集体信念和行为、文化积累及集体智能,指出人类学习社会、空间与非社会拓扑的潜在的共同机制,并认为当前人工智能可以与脑神经网络及人类社会网络中获得的洞察结合,既加深对人类集体认知的理解,同时促进集体机器智能的发展。本综述回顾的各项研究使用了在图论、统计学、数学、物理学和神经科学中开发的各种跨学科方法。该综述收录于《英国皇家学会哲学通报B》(The Philosophical Transactions of the Royal Society B)2022年1月发布的“集体知识和文化积累在动物、人类及机器中的涌现”主题特刊中。
研究领域:网络拓扑,社会网络,认知神经科学,人工智能

论文题目:
Collective minds: social network topology shapes collective cognition
论文链接:
https://royalsocietypublishing.org/doi/full/10.1098/rstb.2020.0315
目录
摘要
引言
网络拓扑协调集体记忆
网络拓扑塑造信念和规范
网络拓扑塑造集体智能
社会网络拓扑塑造神经反应
存在引导社会及非社会拓扑的普适机制?
网络拓扑应用于集体机器智能
结论:人类与机器的集体认知拓扑
摘要
人类的认知并不是孤立的,而是通过集体学习和记忆塑造而成的。与虫群或兽群不同,人类社会网络具有不同的拓扑,服务于不同的集体认知及行为模式。在此,我们回顾了将网络结构与心理学、神经学实验及模型相结合,以理解社会网络的拓扑如何塑造集体认知的相关研究。首先,我们回顾了基于图论的方法在集体记忆、信念传播和问题处理的行为学实验中的应用。这些研究表明,不同的交流网络拓扑以不同的方式同步或整合知识,并服务于不同的集体目标。第二,我们讨论了神经影像学研究,这些研究表明人类大脑编码了更大的社会网络拓扑,并显现出与朋友及社群关系相似的神经学模式(比如在看电影的时候)。第三,我们讨论了学习社会与非社会拓扑之间的认知相似性(例如在空间和联想学习方面),以及参与处理社会和非社会拓扑的共同脑区。最后,我们讨论了最近的一些机器学习方法,通过多主体人工网络进行集体交流及合作。将网络科学与认知、神经和计算方法相结合,有助于探究社会结构是如何塑造认知的。反之,它也可以帮助设计目标导向的社会网络拓扑。
本文是“集体知识和文化积累在动物、人类及机器中的涌现”主题特刊的一部分。

1. 引言
人类的认知并不是孤立的。从工具的使用、语言和数学到对世界及道德的信念,人类的认知是通过在社会网络中学习和记忆而形成的。与虫群或兽群不同,人类社会网络具有不同的结构,包括强、弱、集群,甚至是等级关系,服务于不同的环境。人类通过这种网络传递并共享信息,以同步其集体记忆、知识和信念,或发现并整合不同的信息和知识。本综述侧重于有关交流网络结构(即社会网络拓扑)如何塑造集体认知的相关实证及计算研究。具体来说,我们想知道社会网络拓扑是通过哪些心理学、神经学及计算学有关学习及记忆的原理,来同步集体记忆和认知的。
尽管在过去的几十年间,已有一些著名论文和书籍讨论了合作和社会网络对集体产出所发挥的作用,但似乎并没有综述专门将社会网络拓扑与心理和神经计算综合起来。因此,本综述将重点关注下列方面的重要研究:网络结构(宏观层面)与心理认知(微观层面)在集体记忆 [1,2]、集体信念和行为 [3-5]、集体发现和文化积累 [6]中的相互作用,社会和非社会导向的大型网络神经科学 [7,8] ,以及在机器智能和多主体系统中交流的涌现 [9-11]。
在讨论本综述的重点之前,过去几十年间各个学科对集体认知的研究值得我们注意。Bratman [12-14]、Davidson [15]、Tuomela [16] 和Gilbert [17,18] 等哲学家开创了共同能动性(shared agency)、集体信念和共同意向性(shared intentionality)的理论。这些哲学研究强调了集体信念和意向的重要性,将社会性的概念与道德和伦理联系了起来。人类学家探究了合作及文化累积的演化过程 [6,19],发展和比较心理学家也关注了灵长类和婴儿的心智理论(theory of mind)、共同意向性及合作 [20-24]。另外值得一提的是有关共同意向性、归因和人类文明演化的相关研究,详见本特刊 [25] 中O'Madagin和Tomasello的论述。其他心理学家结合了建模和实验来研究合作 [26]、基于互惠性(reciprocity)和传递性的群体涌现[27] 以及要满足什么条件,两个人才能在感知决策中比他们中的任意一个表现更好 [28]。对这些文献的全面回顾超出了本文的范围,只有更广泛的回顾才能将现有观点与传统哲学、人类学和决策科学的观点结合起来。
研究人类交流网络的拓扑使我们能够理解、解释、建模并预测大型网络中集体知识的涌现和动力学。在过去数十年里,数学家、物理学家、神经科学家、计算机科学家、社会学家和经济学家的开创性研究,建立了复杂网络科学,这些研究在之前的刊物中已经有精彩的回顾[29-33]。
本文特别关注网络拓扑与计算及认知科学相关方法的结合。对图论的理解,使得我们可以研究交流网络如何与心理学及神经计算相互作用,从而形成集体认知。此外,它还使得我们可以做出目标导向性预测,并设计干预措施,以达到期望的集体认知结果。这种期望的集体性结果可以涵盖下列方面:预测并打击有关全球流行病的错误信念,促进课堂学习的最佳结构,在选举前同步大众记忆,优化致力于快速发现疫苗的科学工作队的连接,关于能增强研究人员的人类集体认知研究,以及设计有效的多主体机器智能。
本文在上述范围内,回顾了人类集体认知的最新研究方向,并在最后简要讨论了多主体机器智能在当前及未来的发展方向。我们回顾了网络拓扑如何影响集体记忆(§2)、集体信念和行为(§3)、文化积累及集体智能(§4)。然后,我们讨论了大脑的神经反射如何捕捉个体社会网络的拓扑(§5),并讨论了社会和非社会拓扑认知中的常见神经学发现(§6)。最后,我们介绍了在多主体机器学习中的应用(§7),总结了人类和机器中的社会网络拓扑(§8)。

图1. 网络拓扑的入门知识。社会及非社会网络可以通过图进行分析。(a)展示了一个集体记忆研究 [2] 的网络拓扑示意图。节点(Node),即社群中的个体,用人形表示。边(Edge),即两个节点(个体)之间的直接联系,用线表示。集群(cluster),包括桥接关系(bridge ties,节点间没有公共连接)和集群关系(cluster ties,的节点之间则存在多条公共关系)也在图中标出。节点的度(degree),即节点的关系数,用绿色标记。不同图结构的标准参数包括:随机性(b)、聚类关系(a,c)、网络直径(最大路径长度)和平均路径长度。著名的图拓扑包括:网格结构图、随机环状图和集群社区结构图(c)。
2. 网络拓扑协调集体记忆
理解集体认知的一个关键问题是,交流网络的结构(图 1)如何协调集体共同记忆和信念。从朋友圈到大型社区,共同的记忆往往会塑造群体认同,反过来又促进了集体行为。需要注意的是,集体记忆在社会学和心理学文献中有着不同的定义。在本文中,集体记忆是指社会网络或社区成员之间记忆的融合[1,2]。但在社会科学中集体记忆还有其它的概念,比如由社会维护的公共符号 [34,35],例如战争纪念碑等。
在一些研究中,Coman、Momennejad及其合作者通过图论、行为学实验、复杂和时间网络分析以及基于主体的仿真等方法,来研究集体记忆 [1,2,36]。为此,他们设计了基于实验室的集体记忆对照实验,在此期间,受试者会被分配到预先编排的虚拟环境交流网络中。受认知神经科学中表征相似性分析(representational)的启发,研究人员设计了新的行为学分析方法,可以在个人意识不到广泛拓扑的情况下,度量社会网络中记忆的融合[1,2]。
每个实验环节都由多名受试者组成(例如10人或16人),每个人面对电脑屏幕,随后通过电脑的文本聊天窗口与预先指定数量的其他受试者进行虚拟的交互。所有实验都按照三个阶段进行设计(图 2):在第一阶段,受试者单独学习相同的材料并进行回忆测试(会话前回忆)。在第二阶段,每名受试者通过一个弹出式聊天窗口,进行一系列二元会话,在此期间他们可以讨论从学习材料中记住的内容。受试者不会知道他们在和谁进行交谈,也不会意识到存在着大型网络结构。在第三阶段,每名受试者再次单独进行回忆测试(会话后回忆)。通过会话前及会话后回忆的行为学方法,我们可以在集群拓扑及非集群拓扑这两种不同的拓扑条件下,度量由于会话导致的记忆融合(图 2)。受神经成像中类似技术的启发,研究人员设计并进行了一系列表征相似性分析(图 2),以计算和比较与网络拓扑相关的个人和集体记忆。
作者首先将第一阶段和第三阶段的行为回忆反应作为回忆项目的向量进行编码。例如,某项研究有16个回忆问题 [1],则可以得到2组16维的向量作为对话前后回忆的结果。如果受试者正确回忆了某个项目,则该项目编码为1,否则为0。然后计算每对受试者的记忆相似度(图2),即计算会话前和会话后阶段的回忆向量之间的相关性或点积。记忆相似度(Mnemonic similarity)衡量了一对受试者回忆和遗忘项目的相似程度。将所有配对的记忆相似度取平均,就能算出该网络的记忆融合度(Mnemonic convergence),即整个网络回忆和遗忘项目的相似度(图2)。将会话前后的记忆相似度相减,就可以得到记忆协调度(Mnemonic alignment),即该对受试者的记忆(回忆和遗忘)在对话后变得更加相似的程度。将所有配对的记忆协调度做平均,就可以度量网络集体记忆。
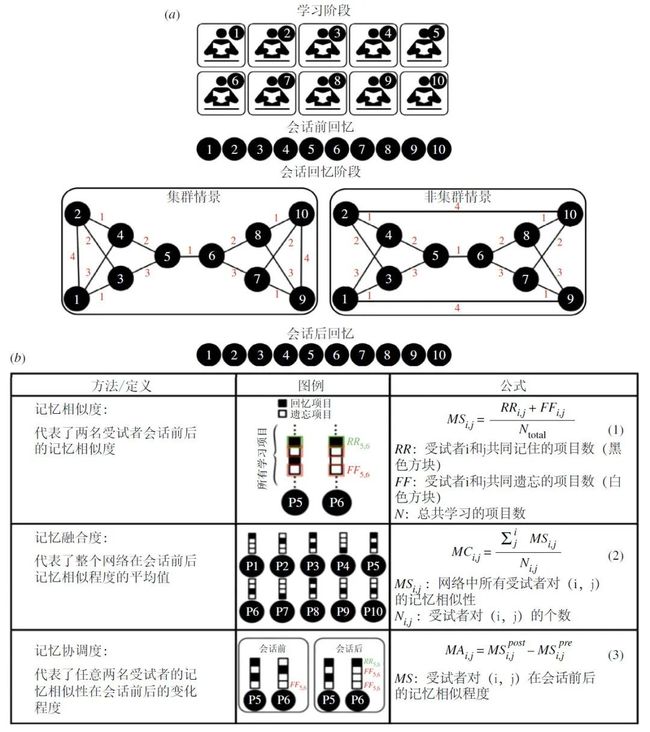
图2. 研究网络拓扑对集体记忆的作用。(a)在实验室形成的交流网络中,度量集体记忆的实验设计 [1,34]。在受试者不知情的情况下,研究者给实验的每个环节都分配了一个10人构成的拓扑。有三个阶段:个体学习并回忆测试阶段、会话阶段、以及会话后个体回忆测试阶段。(b)计算记忆相似程度(随会话而变)的方程。数字代表网络中的成员,例如P6代表6号受试者。
通过上述实验范式(图2),作者研究了网络的结构或拓扑对会话后受试者记忆融合程度(这是集体记忆融合的一个衡量标准[1])的影响。数据是从10名受试者构成的网络中收集的,在该网络中,每名受试者都进行了3次会话,这些会话按照集群或非集群网络拓扑有序进行。作者测量了8个集群的10人网络和8个非集群的10人网络(图2)。
该研究的假说是,这些成对受试者的记忆会根据其分离程度进行协调,直接对话的受试者的协调程度最高,而会话距离最远的受试者的协调程度最低(图 3)。该假设在协调的行为学测量中得到了证实(图 3)。成对的结果还表明,大型网络的拓扑会对集体记忆产生影响。与非集群网络相比,集群拓扑的网络具有更大的网络直径,即网络最远成员间具有更长的路径。结果表明,具有较小直径的网络(非集群拓扑)会比具有集群图结构或拓扑的网络更容易发生记忆融合。
Momennejad及其合作者在另一项实验 [2] 中发现,在一个固定的交流网络拓扑中,交流随时间逐渐展开的顺序,决定了记忆的协调程度。也就是说,他们发现,具有弱或桥接关系的个体间先进行信息交换,会加剧集体记忆的融合,这与Granovetter对于弱关系强度的观点相符 [37]。这些个体有直接的联系,但与其他个体没有任何共同的联系。(要直观地了解他们的网络状态,可以用一个类比:他们是朋友,有交流,但他们的朋友不是朋友,也不互相交流。)
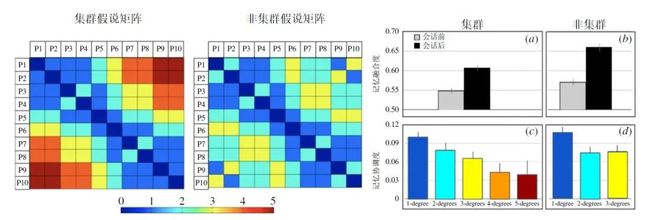
图3. 网络拓扑对集体记忆的协调作用。该图显示了图2中描述的集群拓扑条件及非集群拓扑条件所对应的记忆相似度假设矩阵。数字代表网络中的成员,例如P6代表6号受试者。相似度范围从0(自身;深蓝色)到5(最大分离度;深红色)。行为学结果表明,平均而言,非集群网络(b)中的会话后记忆协调度比集群网络(b)更高。这一发现可以用记忆相似度假设矩阵来解释——记忆的协调程度取决于个体的分离程度。集群网络拓扑具有较大的分离度(即较长的节点间距或网络直径),会导致更低的融合性。个体间的协调程度取决于其在社会网络中彼此的分离程度(c,d)。
上述研究表明,交流网络的拓扑和时间顺序,可以决定网络中个体之间的记忆协调度,即使在他们无直接交互的情况下。在最近的一项计算研究 [36] 中,我们将多主体或基于主体的模拟,与记忆心理学中的提取诱发遗忘模型相结合 [38,39],并成功模拟了这些有关集体记忆的行为学发现。总的来说,这些行为学和计算学研究提供了一种定量方法,可以根据网络拓扑及记忆遗忘原理,测量在集体水平或介观尺度现象的涌现。
这些最新进展,为研究人类认知及行为在微观、介观和宏观尺度之间联系的理论及实验方法开辟了道路。集体实验的图论方法,以及基于实验的带参数的机构模拟[40],可以帮助理解、预测和比较人类网络的不同拓扑的行为。
3. 网络拓扑塑造信念和规范
集体记忆通常构成共同信念体系的基础,而信念则决定了我们记忆的方式[41]。宗教、政治选择及健康决策,是共同信念及其动态变化如何引导个体及集体决策的重要事例。计算模拟已经确定了信念在社会网络中传播的机理 [42]。最近的经验证据表明,集体信念可以根据社会网络的拓扑进行同步 [43]。
Vlasceanu和Coman进行了图2所示研究的信念同步版本 [1,5] 。他们发现,与集体记忆的对应研究 [1] 类似,社会网络的拓扑也会影响信念的协调程度。这种信念协调,甚至会发生于无直接交流的个体之间,且个体间信念相似度的变化程度与其在网络中的分离程度相关。
作者还在由12人构成的实验网络中,研究了公共演讲者为某信念的背书如何影响信念的协调[43]。在实验过程中,受试者首先会评估一组信念陈述的可信度,然后完成一项干扰任务(控制条件)或听取认可某些信念的公共演讲(处理条件)。作者发现,公共演讲改变了一些信念的记忆可及性,而其又会影响网络交流并被其增强,导致后续的信念同步化。未来的研究需要着眼于,个体的社会网络对一个公共演讲者的喜爱,如何塑造其影响的方向。
本文侧重于社会网络拓扑对集体认知的作用,而不涉及广义的行为。然而,有关健康、教育和政治的信念支配着大尺度的人类集体行为模式。因此,社会网络拓扑塑造了行为,这一假说是合理的。Centola在一系列开创性的研究中,为这一假说提供了证据,建立了网络拓扑和集体行为之间的联系 [3,4]。他们调查了健康行为通过社会网络传播的机制,并确定了哪些拓扑使得这些行为的传播更为有效。他们发现,如果个体在更广泛的社会网络中,受到大量关系密切者对该健康行为的信息强化,他们会更倾向于采取该健康行为。因此,对健康行为的采用,在集群规则网络拓扑中,会比在随机网络中更有效地进行传播 [3]。在另外一项研究中,Centola调查了社区的组成如何影响健康行为的采用。他们发现,两名个体间的同质性(即两者社会关系的相近程度)会增加他们对彼此行为的影响。然而,这种模式更广泛的后果是,大型集群网络中那些不太健康的个体,更可能建立密切的联系并相互影响,从而减少了他们与更健康的个体互动并受其影响的可能性 [4]。
虽然本文是对网络拓扑在集体认知中的作用的综述,着眼于认知神经科学和机器学习,但最近的一系列论文,就社会网络拓扑如何影响健康行为、资源分配和资源不平等,以及集体智能(如观察到群体的平均反应比个人反应更接近真值)等问题进行了讨论[34-39]。虽然这些都是重要且引人入胜的相关话题,但已经超出了集体认知中微观-宏观相互作用的范畴。不过,一些利用网络方法改变社会规范的论文值得特别注意。Banerjee及其合作者研究了小额信贷在农村中的传播,结果表明,如果首先将信息提供给中心个体(通过其在社会网络中特征向量的中心性来度量),则被采纳的信息会在社区中更有效地进行传播 [44]。在另一项影响深远的研究中,Paluck及其合作者对24000多名学生进行了反冲突干预。结果表明,个体会更关注社会网络中的“社会参照”角色(有影响力的个体),而针对社会参照个体进行干预,使得1年内发生的冲突减少了30%,比普通干预更为有效。此后,他们又进一步研究了工厂的社会变化:规范变更、劝说方式和工厂工人们对权威和正义的态度随时间和基于网络的进步 [46-49]。
总而言之,这些研究阐明了网络拓扑在信念、规范及行为在网络中的传播机制的作用,以及集中式干预(如让集体暴露于公众的认可或高影响力个体的认可)如何改变集体信念和规范。上述研究,连同集体记忆及信念形成的相关结果,展现出网络拓扑在在大尺度网络中打击错误信息(如有关健康行为)或有害的两极分化等方面的重要潜能。通过对错误信息网络建立网络拓扑,我们可以设计一系列干预工具,用于在危机时期同步大型社群中的记忆及信念。这些工具可以影响社会网络的拓扑,甚至可能针对桥接关系及孤立集群(信息茧房)进行干预和集中式公开演讲。
4. 网络拓扑塑造集体智能
想想你的智能手机、布鲁克林大桥及国际空间站。实现这些复杂技术中的任何一个,都需要几个世代的技术积累,可见解决这些问题能力的水平总是远远超出个人的智能水平。有人提出,取得累积性文化成就(如复杂技术等)的能力,取决于协作网络的规模及连通性 [6]。
在一项开创性研究中,Derex和Boyd [6]探究了协作网络的拓扑及时间顺序如何带来创造。他们研究了一个由6名受试者构成的协作网络,尝试在虚拟情景中探索一种具有三级结构的化合物。在第一种情景中,协作网络中的6名受试者一起工作(全连接),而在第二种情景中,他们在3个2人小组中工作,并在整个实验过程中进行了2次成员交换(部分连接)。结果表明,与全连接情景相比,部分连接情景下的受试者得到了更多层次的发明。
因此,正如前面所讨论的那样,虽然全连接可能有助于记忆及信念的融合,但部分连接可能会增加文化积累,这是复杂和累积性创造所需要的。这些发现为本文的核心论点提供了有力的支持:特定的社会网络拓扑可以服务于特定的集体结果需求。基于该假设的后续研究,可以为干预设计带来启示:对网络特定部分设计目标导向的结构或进行针对性的干预,来实现预期的结果。这些预期结果的实例包括预测并打击有关全球大流行的错误信息,以及设计科研网络的最优交流以实现快速疫苗开发。
Wooley及其合作者将集体智能定义为,超越集体中大部分个体,集体解决各种任务和问题的能力 [50,51]。他们通过多项研究,探究了团队的组成及多样性如何增强其可度量的集体智能,并帮助得到特定问题的最优解 [50,51]。他们发现,在从眼睛照片中检测情绪的社会智力测试中,团队成员的社会敏感度可以预测团队的集体智能 [52]。然而,这种社会敏感度,并不能度量个体对社会网络中更广泛关联拓扑编码的敏感度。
上述Derex与Boyd的研究为“网络的拓扑及连通性可以影响社群集体智能”的假设提供了初步证据。还有研究在社会网络中建立了协作学习 [53],并探究了团队规模及组成在复杂任务中的作用 [54]。诚如我们所见,不同的网络拓扑为不同的目标提供了特定的最优解决方案。一项有待后续研究的假说是,协作网络的集体智能,可能取决于网络在重构其协作连接以适应不同任务的需求方面的灵活性。
另一个有待后续研究的方向,是探究集体智能是否仅取决于对社会信号的敏感度,还是说,它也与社群成员感知网络中其他人的状态,及社会网络的广泛拓扑的能力有关。这些研究可以受益于当前神经影像学研究中开发的方法(会在下一节中进行讨论),后者发现,一个人的大脑会自发地对社群成员的社会网络状态进行编码。
5. 社会网络拓扑塑造神经反应
上述章节回顾了一些行为学证据,证明人类的记忆与信念会变得与自己的社群成员相似,而且这种相似的程度会体现个体与社群其他成员的测地距离,或者说分离程度。鉴于这些行为学发现,我们会自然而然得出一种假说:社会网络拓扑也会影响社群成员间大脑信号的相似性。最近有项研究探究了人类大脑是否对其他个体的网络状态敏感 [7]。作者推断,人类社群与虫群或兽群不同,由多样化的联系及结构组成,因此人类的大脑很可能已经演化到可以承载引导复杂社会网络的认知需求了。这意味着人类可以追踪直接(单级连接)、间接或更遥远(多级连接)的社会关系——延伸到对于更广泛的社会网络拓扑的理解。
该研究通过特征向量中心性等图论方法,对同一学术群体中的275名学生进行了分析,并对其中21名成员进行了功能性磁共振成像(fMRI)。特征向量中心性度量了一个成员的影响力大小,及其连接的有影响力的个体的多少(这与图1中的中心度,仅度量一个成员连接的多少,是不同的)。在研究过程中,每个受试者都看了与其具有不同分离度个体的照片。受试者包括了有特征向量中心度(作为影响力的度量标准)的个体,也包括了那些在群体中原本不相关个体间成为桥梁纽带(作为间介性的度量标准)的个体(图1)。神经模式分析表明,当个体看到其它群体成员的照片时,他们的大脑会激活正在观看的这个人的网络位置。值得注意的是,这种对群体成员特定网络位点的神经反应是在大脑中自发激活的,即在没有明确的目标需要这种知识的情况下。这与之前的假设相一致,即人类大脑可能已经适应了对社会网络关系进行拓扑编码,而不是仅仅对个体的直接关系编码 [55]。
一系列研究通过fMRI扫描,对社群成员在观看视频时大脑激活的相似性进行了探究。研究者绘制了个体的社会网络图,根据不同群体成员的不同中心性,度量了其网络状态,并分析了大脑对视频反应的相似性与社会网络紧密度之间的关系。其中一项研究 [56] 发现,个体在观看视频时的神经反应与其相近的社群关系者更相似。这种神经相似性的程度,反应了对象在社会网络中的距离:分离程度较小的朋友间的大脑反应更相似,而分离程度较大的社群成员间的反应则不太相似。另一项研究表明,社区成员在观看视频时,大脑活动会变得与网络中高特征向量中心性(高影响力)的个体更相似,作者称之为“神经影响力" [57]。
至此,我们已经回顾了一些图论研究:社会网络中集体记忆 [1,2] 、行为与信念 [43] 的同步;社群成员间的网络相似性的协调 [56,57];以及社群成员有关网络位点的神经编码 [7]。这些研究表明,人类在社会及非社会认知中对网络结构具有更为广泛的学习能力。
6. 存在引导社会及非社会拓扑的
普适机制?
人类的社会群体,比我们的演化近亲大得多,且研究表明,社会网络的大小与灵长类 [55] 及其他哺乳类动物 [58] 的脑容量相对应。与虫群和兽群不同,人类社群由具有不同结构的社会网络组成。本特刊中的一些论文侧重于累积文化的演化及人口结构,并将其作为从温饱转向集体智能的基础,而其它一些论文则关注狩猎-采集者的社会网络,以了解文化的演化过程(见 [59,60],以及 [61])。虽然已经有跨物种的社会学习的新兴研究关注文化演化和集体知识(详见 Garland 等人 [62],Gruber 等人 [63],Whiten 等人 [64],Wild 等人 [65],以及在本特刊中Williams及Lachlan [66] 的讨论),本文特别关注网络拓扑对人类集体认知的作用,故在演化框架方面不作过多介绍。也就是说,可以合理地假设,人类大脑已经演化到可以承载引导复杂社会网络的认知需求;反之,适应社会网络增长的需求,也许促成了更广泛的学习与引导复杂网络的实践与文化。虽然尚不清楚在社会、空间或关联环境中,神经和文化进行图学习的能力,是如何产生并共同演化的,以及哪种能力在先,但在整个结构学习研究中考虑这些共同工具和结论,是很有帮助的 [8]。
一系列有关联想、统计和表征学习的行为学、神经成像以及计算学研究,确认了人类具有学习多步拓扑及序列的社群结构的能力 [67-69]。连接主义者(connectionist)及强化学习的计算模型框架,都提供了大脑如何将序列结构的联想学习推广到对更大结构的学习的解释 [8]。最近的行为学和神经科学研究,已经确认了社会结构学习中的计算学习原理 [70] ,以及这种学习所依赖的脑神经网络 [68,71,72]。
接下来的一系列研究,描绘了人类学习和引导非社会网络结构能力的更广阔图景。研究表明,序列的统计学习是人类学习和表示图和网络的基础,无论是对于8个月大的儿童[73],还是在对高阶时间结构的关联学习中提取时间事件的统计数据时 [74],并发现类似的原理可以被推广到语言习得领域 [75,76]。还有人研究了在社会及非社会结构中学习的个体差异 [77],以及对局部模式的学习如何引起对网络拓扑的学习 [78,79] 。
最近,Schapiro 及其合作者发现,人类在接受一系列的个体刺激时,会隐式地对更大的网络结构进行学习。通过fMRI,他们表明,这种图学习过程体现在前额叶 (PFC) 和内侧颞叶等脑区中 [67]。这种范式已被其他研究者用来研究人类是如何学习具有不同拓扑的统计结构的[80]。此外,一系列行为学及神经影像学的研究已经使用了强化学习框架,提供了学习多步关联的关系结构(也称后继表征)的证据 [68,69,71,81],这些证据可能也解释了大脑如何学习社会网络的拓扑。目前人类神经影像学支持这样的观点:新的社会等级推断,依赖于与导航类似的神经机制。
社会拓扑学习与空间拓扑导航、捷径探索及学习非空间关联拓扑之间的相似性,呼唤我们对它们的共同机制进行更多的比较研究。两个涉及社会、空间及其他拓扑模式学习的特定脑区,是内侧前额叶(mPFC)和海马-内嗅皮层复合体 [72,81-84]。在这些脑区中,内侧前额叶在人类和非人动物间的差异最大,且其前部与社会认知特别相关。内侧前额叶前部(布罗德曼10区),在处理不同的持续性任务,以及进行多任务 [85-87]、类比推理 [88] 和社会推理 [89-91] 时,也涉及表征前瞻性任务。
需要注意的是,这些研究在很大程度上借鉴了神经科学、数学、图论和物理学等领域的各种专业知识及工具 [92]。通过识别人类认识社会及非社会拓扑的能力背后的共同的脑神经网络,可以为理解集体认知及文化演化提供新的启发。跨学科实验及建模有助于阐明人类大脑学习社会及非社会网络的能力共同演化的动态过程。反之,理解人类及非人灵长类大脑的学习图和拓扑的神经计算能力,也有助于开发多主体架构的集体机器智能。
7. 网络拓扑应用于集体机器智能
当代机器学习一个蓬勃发展的方向是多主体学习及协作式人工智能(AI)。这些方向的研究及创新涵盖了AI-AI交互及AI-人交互(包括基于自然语言处理的交流 [93] ),以及构建AI工具,用于增强人-人交互 [94] 等多种方面。本特刊也包括了许多这些方向的研究,例如基于集体社会机器人的人工文化实验(见 [95]),基于蜂群机器人的社会学习的具身演化的研究(见[96])以及对机器人身体及文化学习的人工演化的探究(见[97])。目前的研究范围从玩计算机游戏(如Project Malmo、Xbox games、我的世界 [9,98] 和星际争霸 [11,99])的深度增强学习智能体,发展到了多主体网络 ,这些网络演化出了基于交流的社会影响力[10]、基于交互的学习 [100] 以及交互式的元学习 [101]。智能体交互图已被用于评估多主体系统中的泛化 [102]。多主体机器学习的未来发展方向可以与从脑神经网络及人类社会网络中获得的洞察相结合,既加深对人类集体认知的理解,又促进集体机器智能的发展。
一个前瞻性的计算学研究方向是,在一个多主体系统中,比较不同独立个体神经架构所涌现的不同社会结构。可以设想至少两个相关的研究方向。首先,可以探究最优的网络拓扑如何在多主体环境中涌现,以满足特定的任务需求或目标。这类系统的目的从协作、竞争(例如Xbox games)到辅助人类各不相同。其次,可以研究将网络架构演化的原理与多主体问题情景相结合,来深入理解神经架构(在独立个体中)与社会及生态多主体网络拓扑的共同演化过程。鉴于图学习领域的最新进展 [103],这些方向都非常应景。
这些方向的研究进展,可以为个体的脑神经架构与物种在大型复杂社会、空间及环境网络中导航需求的对应关系与共同演化,提供理论支持。哪些社会网络结构源于有着不同目标的多主体系统?哪种神经网络或大脑架构可以承载特定物种的多主体行为?这些研究方向,为生物学及人工智能范畴下,神经网络与社会网络如何共同演化的探索,描绘了令人激动的前景。
8. 结论:人类与机器的集体认知拓扑
人类的大脑及文化内嵌在广泛的社会及生态系统中。与虫群及兽群不同,人类的社会网络有着不同的成分和拓扑。在此,我们回顾了支持“社会网络拓扑塑造集体认知及行为”这一假说的相关研究。我们主要关注于基于微观及宏观相互作用的心理学及神经科学研究。
综合这些研究可以发现:(1) 人类的记忆、信念及行为会与他们有社会关系的人,以及他们从未直接接触的社群成员同步 [1–5];(2) 人类大脑会自发地处理其他个体在自己社会环境中的网络状态,而观看视频时大脑反应的相似性预测了一个组群内的友谊关系 [7,56];(3) 大脑编码比直接关系更广泛的网络拓扑的能力,反映了大脑学习非社会拓扑及认知图的能力 [104]。我们还回顾了神经科学中有关学习及记忆的一些证据,指出了学习社会、空间与非社会拓扑的潜在的共同机制 [8,67,77];(4) 最近的一些深度学习算法则在多主体机器学习中,将上述研究与集体认知联系了起来 [10]。这些多样化的研究支持了一个假说——大脑获取并引导复杂大型神经网络拓扑的能力,可能是与人类不断增长的网络规模及逐渐多元化的社会拓扑共同演化的。而有关社交及情感障碍的研究可以阐明社会与非社会图学习间的异同。
本综述所回顾的各项研究,使用了在图论、统计学、数学、物理学和神经科学中开发的各种跨学科方法,对复杂网络中的社群结构进行了分类及表征 [105,106],对动态复杂系统进行了时间网络分析(例如,在集体记忆的相关研究中,分析对话顺序的作用)[2],用多项目记忆向量的相关性来进行表征相似性分析,来比较适应后的多维向量(例如分析神经模式 [84]),从而比较行为中的记忆融合 [1,34]。
用于分析复杂网络架构的图论工具,同样适用于大脑及社会网络。正如不同架构的神经网络以不同的方式塑造并整合了信息,特定结构或拓扑的社会网络会通过不同的方式同步或整合知识。后续研究可以结合实验及计算的方法,来研究那些能够处理大型社会网络拓扑的神经网络的共同演化,以及那些服务于不同集体功能和结果的拓扑的涌现。
将实验与计算方法相结合,研究者得以探究社会网络拓扑是如何塑造集体认知及行为的,更好地设计目标导向的社会网络拓扑,以实现预期的结果(例如纠正有关全球大流行的错误信息或是协调疫苗的快速开发)。跨物种的社会及非社会图学习的比较研究,可以加深对人类、其它物种及机器的神经和社会机制的演化过程的理解。这种跨学科的研究方法,也使得研究者可以在人类集体认知相关知识的启发下,设计更有效的多主体机器智能。反之,基于集体智能的多主体机器学习模型,有助于对“复杂神经架构与复杂社会网络共同演化”这一假说进行理论化及测试。
参考文献
[1] Coman A, Momennejad I, Drach RD, Geana A. 2016Mnemonic convergence in social networks: the emergent properties of cognition at a collective level. Proc. Natl Acad. Sci. USA 113, 8171-8176.
[2] Momennejad I, Duker A, Coman A. 2019Bridge ties bind collective memories. Nat. Commun. 10, 1578.
[3] Centola D. 2010The spread of behavior in an online social network experiment. Science 329, 1194-1197.
[4] Centola D. 2011An experimental study of homophily in the adoption of health behavior. Science 334, 1269-1272.
[5] Vlasceanu M, Coman A. 2020Network structure impacts the synchronization of collective beliefs. PsyArXiv.
[6] Derex M, Boyd R. 2016Partial connectivity increases cultural accumulation within groups. Proc. Natl Acad. Sci. USA 113, 2982-2987.
[7] Parkinson C, Kleinbaum AM, Wheatley T. 2017Spontaneous neural encoding of social network position. Nat. Hum. Behav. 1, 1-7.
[8] Momennejad I. 2020Learning structures: predictive representations, replay, and generalization. Curr. Opin. Behav. Sci. 32, 155-166.
[9] Perez-Liebana D, Hofmann K, Mohanty SP, Kuno NS, Kramer A, Devlin S, Gaina RD, Ionita D.2019The multi-agent reinforcement learning in MalmÖ (MARLÖ) competition. ArXiv, abs/1901.08129.
[10] Jaques N, Lazaridou A, Hughes E.2019Social influence as intrinsic motivation for multi-agent deep reinforcement learning. In Proc. 36th Int. Conf. on Machine Learning (ICML), Long Beach, CA, PMLR 97.
[11] Vinyals Oet al.. 2019Grandmaster level in StarCraft II using multi-agent reinforcement learning. Nature 575, 350-354.
[12] Bratman ME. 1993Shared intention. Ethics 104, 97-113.
[13] Bratman ME. 2013Shared agency: a planning theory of acting together. Oxford, UK: Oxford University Press.
[14] Bratman ME. 2015Précis of shared agency: a planning theory of acting together. J. Soc. Ontol. 1, 1-5.
[15] Davidson D. 1992The second person. Midwest Stud. Phil. 17, 255-267.
[16] Tuomela R. 1995The importance of us: a philosophical study of basic social notions. Palo Alto, CA: Stanford University Press.
[17] Gilbert M. 1987Modelling collective belief. Synthese 73, 185-204.
[18] Gilbert M. 1989On social facts. Oxford, UK: Routledge.
[19] Boyd R, Richerson PJ. 2009Culture and the evolution of human cooperation. Phil. Trans. R. Soc. B 364, 3281-3288.
[20] Tomasello M, Melis AP, Tennie C, Wyman E, Herrmann E. 2012The interdependence hypothesis. Curr. Anthropol. 53, 673-692.
[21] Tomasello M. 2009The cultural origins of human cognition. Cambridge, MA: Harvard University Press. Crossref.
[22] Tomasello M, Carpenter M, Call J, Behne T, Moll H. 2005Understanding and sharing intentions: the origins of cultural cognition. Behav. Brain Sci. 28, 675-691; discussion 691–735.
[23] Zahavi D, Satne G. 2015Varieties of shared intentionality: Tomasello and classical phenomenology. In Beyond the analytic-continental divide (eds Bell JA, Cutrofello A, Livingstone PM), pp. 313-334. Oxford, UK: Routledge.
[24] Tomasello M, Carpenter M. 2007Shared intentionality. Dev. Sci. 10, 121-125.
[25] O'Madagain C, Tomasello M. 2021Shared intentionality, reason-giving and the evolution of human culture. Phil. Trans. R. Soc. B 377, 20200320.
[26] Peysakhovich A, Nowak MA, Rand DG. 2014Humans display a ‘cooperative phenotype’ that is domain general and temporally stable. Nat. Commun. 5, 4939.
[27] Gray K, Rand DG, Ert E, Lewis K, Hershman S, Norton MI. 2014The emergence of ‘Us and Them’ in 80 lines of code: modeling group genesis in homogeneous populations. Psychol. Sci. 25, 982-990.
[28] Bahrami B, Olsen K, Latham PE, Roepstorff A, Rees G, Frith CD.2010Optimally interacting minds. Science 329, 1081-1085.
[29] Barabási AL. 2003Linked: the new science of networks. Am. J. Phys. 71, 409-410.
[30] Watts DJ, Strogatz SH. 1998Collective dynamics of ‘small-world’ networks. Nature 393, 440-442.
[31] David E, Jon K. 2010Networks, crowds, and markets: reasoning about a highly connected world. New York, NY: Cambridge University Press.
[32] Barabási AL. 2013Network science. Phil. Trans. A Math. Phys. Eng. Sci. 371, 20120375.
[33] Watts DJ. 2004Six degrees: the science of a connected Age. New York, NY: W. W. Norton & Company.
[34] Hirst W, Yamashiro JK, Coman A. 2018Collective memory from a psychological perspective. Trends Cogn. Sci. 22, 438-451.
[35] Olick JK, Vinitzky-Seroussi V, Levy D. 2011The collective memory reader. New York, NY: Oxford University Press.
[36] Sievers B, Momennejad I. 2019SAMPL: the spreading activation and memory plasticity model. bioRxiv, 778563.
[37] Granovetter MS. 1973The strength of weak ties. Am. J. Sociol. 78, 1360-1380.
[38] Anderson MC, McCulloch KC. 1999Integration as a general boundary condition on retrieval-induced forgetting. J. Exp. Psychol. Learn. Mem. Cogn. 25, 608-629.
[39] Norman KA, Newman EL, Detre G. 2007A neural network model of retrieval-induced forgetting. Psychol. Rev. 114, 887-953.
[40] Momennejad I, Sinclair S, Cikara M. 2019Computational justice: simulating structural bias and interventions. bioRxiv.
[41] Schacter DL, Scarry E. 2001Memory, brain, and belief. Cambridge, MA: Harvard University Press.
[42] Friedkin NE, Proskurnikov AV, Tempo R, Parsegov SE. 2016Network science on belief system dynamics under logic constraints. Science 354, 321-326.
[43] Vlasceanu M, Morais MJ, Duker A, Coman A. 2020The synchronization of collective beliefs: from dyadic interactions to network convergence. J. Exp. Psychol. Appl. 26, 453-464.
[44] Banerjee A, Chandrasekhar AG, Duflo E, Jackson MO. 2013The diffusion of microfinance. Science 341, 1236498.
[45] Paluck EL, Shepherd H, Aronow PM. 2016Changing climates of conflict: a social network experiment in 56 schools. Proc. Natl Acad. Sci. USA 113, 566-571.
[46] Prentice D, Paluck EL. 2020Engineering social change using social norms: lessons from the study of collective action. Curr. Opin. Psychol. 35, 138-142.
[47] Wu SJ, Paluck EL. 2020Participatory practices at work change attitudes and behavior toward societal authority and justice. Nat. Commun. 11, 2633.
[48] Wu SJ, Paluck EL. 2021Designing nudges for the context: golden coin decals nudge workplace behavior in China. Organ. Behav. Hum. Decis. Process 163, 43-50.
[49] Wu SJ, Paluck EL. 2021Having a voice in your group: increasing productivity through group participation. psyarxiv.com.
[50] Woolley AW, Chabris CF, Pentland A, Hashmi N, Malone TW. 2010Evidence for a collective intelligence factor in the performance of human groups. Science 330, 686-688.
[51] Woolley AW, Aggarwal I.2015Collective intelligence and group performance. Curr. Dir. Psychol. Sci. 24, 420-424.
[52] Megías-Robles A, Gutiérrez-Cobo MJ, Cabello R, Gómez-Leal R, Baron-Cohen S, Fernández-Berrocal P. 2020The ‘Reading the mind in the Eyes' test and emotional intelligence. R. Soc. Open Sci. 7, 201305.
[53] Mason W, Watts DJ. 2012Collaborative learning in networks. Proc. Natl Acad. Sci. USA 109, 764-769.
[54] Mao A, Mason W, Suri S, Watts DJ. 2016An experimental study of team size and performance on a complex task. PLoS ONE 11, e0153048.
[55] Dunbar RIM, Shultz S. 2007Evolution in the social brain. Science 317, 1344-1347.
[56] Parkinson C, Kleinbaum AM, Wheatley T. 2018Similar neural responses predict friendship. Nat. Commun. 9, 332.
[57] Sievers B, Welker C, Hasson U, Kleinbaum AM, Wheatley T. 2020How consensus-building conversation changes our minds and aligns our brains. PsyArXiv.
[58] Pérez-Barbería FJ, Shultz S, Dunbar RIM. 2007Evidence for coevolution of sociality and relative brain size in three orders of mammals. Evolution 61, 2811-2821.
[59] Kirby S, Tamariz M.2021Cumulative cultural evolution, population structure, and the origin of combinatoriality in human language. Phil. Trans. R. Soc. B 377, 20200319.
[60] Romano V, Lozano S, Lopez de Pablo J. 2021Reconstructing social networks of Late Glacial and Holocene hunter-gatherers to understand cultural evolution. Phil. Trans. R. Soc. B 377, 20200318.
[61] Migliano A, Vinicius L. 2021The origins of human cumulative culture: from the foraging niche to collective intelligence. Phil. Trans. R. Soc. B 377, 20200317.
[62] Garland EC, Garrigue C, Noad MJ. 2021When does cultural evolution become cumulative culture? A case study of humpback whale song. Phil. Trans. R. Soc. B 377, 20200313.
[63] Gruber T, Chimento M, Aplin LM, Biro D. 2021Efficiency fosters cumulative culture across species. Phil. Trans. R. Soc. B 377, 20200308.
[64] Whiten A, Harrison RA, McGuigan N, Vale GL, Watson SK. 2021Collective knowledge and the dynamics of culture in chimpanzees. Phil. Trans. R. Soc. B 377, 20200321.
[65] Wild S, Chimento M, McMahon K, Farine DR, Shelden BC, Aplin LM. 2021Complex foraging behaviours in wild birds emerge from social learning and recombination of components. Phil. Trans. R. Soc. B 377, 20200307.
[66] Williams H, Lachlan R. 2021Evidence for cumulative cultural evolution in bird song. Phil. Trans. R. Soc. B 377, 20200322.
[67] Schapiro AC, Rogers TT, Cordova NI, Turk-Browne NB, Botvinick MM. 2013Neural representations of events arise from temporal community structure. Nat. Neurosci. 16, 486-492.
[68] Momennejad I, Russek EM, Cheong JH, Botvinick MM, Daw ND, Gershman SJ. 2017The successor representation in human reinforcement learning. Nat. Hum. Behav. 1, 680-692.
[69] Garvert MM, Dolan RJ, Behrens TE. 2017A map of abstract relational knowledge in the human hippocampal-entorhinal cortex. Elife 6, e17086.
[70] Gershman SJ, Cikara M. 2020Social-structure learning. Curr. Dir. Psychol. Sci. 29, 460-466.
[71] Russek EM, Momennejad I, Botvinick MM, Gershman SJ, Daw ND. 2017Predictive representations can link model-based reinforcement learning to model-free mechanisms. PLoS Comput. Biol. 13, e1005768.
[72] Schapiro AC, Turk-Browne NB, Botvinick MM, Norman KA. 2017Complementary learning systems within the hippocampus: a neural network modelling approach to reconciling episodic memory with statistical learning. Phil. Trans. R. Soc. B 372, 20160049.
[73] Saffran JR, Aslin RN, Newport EL. 1996Statistical learning by 8-month-old infants. Science 274, 1926-1928.
[74] Fiser J, Aslin RN. 2002Statistical learning of higher-order temporal structure from visual shape sequences. J. Exp. Psychol. Learn. Mem. Cogn. 28, 458-467.
[75] Romberg AR, Saffran JR. 2010Statistical learning and language acquisition. Wiley Interdiscip. Rev. Cogn. Sci. 1, 906-914.
[76] Friederici AD. 2005Neurophysiological markers of early language acquisition: from syllables to sentences. Trends Cogn. Sci. 9, 481-488.
[77] Tompson SH, Kahn AE, Falk EB, Vettel JM, Bassett DS. 2019Individual differences in learning social and nonsocial network structures. J. Exp. Psychol. Learn. Mem. Cogn. 45, 253-271.
[78] Karuza EA, Thompson-Schill SL, Bassett DS. 2016Local patterns to global architectures: influences of network topology on human learning. Trends Cogn. Sci. 20, 629-640.
[79] Karuza EA, Kahn AE, Bassett DS. 2019Human sensitivity to community structure is robust to topological variation. Complexity 2019, 8379321.
[80] Lynn CW, Bassett DS. 2020How humans learn and represent networks. Proc. Natl Acad. Sci. USA 117, 29 407-29 415.
[81] Stachenfeld KL, Botvinick MM, Gershman SJ. 2017The hippocampus as a predictive map. Nat. Neurosci. 20, 1643-1653.
[82] Chen J, Hasson U, Honey CJ. 2015Processing timescales as an organizing principle for primate cortex. Neuron 88, 244-246.
[83] Brunec IK, Momennejad I.2019Predictive representations in hippocampal and prefrontal hierarchies. BioRxiv.
[84] Momennejad I, Otto AR, Daw ND, Norman KA.2018Offline replay supports planning in human reinforcement learning. Elife 7, e32548.
[85] Momennejad I, Haynes JD.2012Human anterior prefrontal cortex encodes the ‘what’ and ‘when'of future intentions. Neuroimage 61, 139-148.
[86] Momennejad I, Haynes JD. 2013Encoding of prospective tasks in the human prefrontal cortex under varying task loads. J. Neurosci. 33, 17 342-17 349.
[87] Burgess PW, Dumontheil I, Gilbert SJ, Okuda J, Schölvinck ML, Simons JS.2008On the role of rostral prefrontal cortex (area 10) in prospective memory. In Prospective memory: cognitive, neuroscience, developmental, and applied perspectives, (ed. Kliegel M), pp. 235-260. New York, NY: Taylor & Francis Group/Lawrence Erlbaum Associates.
[88] Bunge SA, Wendelken C, Badre D, Wagner AD. 2005Analogical reasoning and prefrontal cortex: evidence for separable retrieval and integration mechanisms. Cereb. Cortex 15, 239-249.
[89] Gilbert SJ, Burgess PW. 2008Social and nonsocial functions of rostral prefrontal cortex: implications for education. Mind Brain Educ. 2, 148-156.
[90] Roca M, Torralva T, Gleichgerrcht E, Woolgar A, Thompson R, Duncan J, Manes F. 2011The role of Area 10 (BA10) in human multitasking and in social cognition: a lesion study. Neuropsychologia 49, 3525-3531.
[91] Gilbert SJ, Williamson IDM, Dumontheil I, Simons JS, Frith CD, Burgess PW. 2007Distinct regions of medial rostral prefrontal cortex supporting social and nonsocial functions. Soc. Cogn. Affect. Neurosci. 2, 217-226.
[92] Adamic LA, Lukose RM, Puniyani AR, Huberman BA. 2001Search in power-law networks. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 64, 046135.
[93] Allison F, Luger E, Hofmann K. 2018How players speak to an intelligent game character using natural language messages. Trans. Digit. Games Res. Assoc. 4, 1-49.
[94] Dafoe A, Bachrach Y, Hadfield G, Horvitz E, Larson K, Graepel T. 2021Cooperative AI: machines must learn to find common ground. Nature 593, 33-36.
[95] Winfield A, Blackmore S. 2021Experiments in artificial culture: from noisy imitation to storytelling robots. Phil. Trans. R. Soc. B 377, 20200323.
[96] Bredeche N, Fontbonne N. 2021A gentle introduction to social learning in swarm robotics. Phil. Trans. R. Soc. B 377, 20200309.
[97] Hart E, Le Goff L. 2021Artificial evolution of robot bodies and control: on the interaction between evolution, learning and culture. Phil. Trans. R. Soc. B 377, 20210117.
[98] Devlin S, Georgescu R, Momennejad I, Rzepecki J, Zuniga E, Costello G, Leroy G, Shaw A, Hofmann K.2021Navigation Turing Test (NTT): learning to evaluate human-like navigation. arXiv.
[99] Arulkumaran K, Cully A, Togelius J.2019AlphaStar: an evolutionary computation perspective. In Proc. of the Genetic and Evolutionary Computation Conf. Companion, pp. 314-315. New York, NY: Association for Computing Machinery.
[100] Xie T, Langford J, Mineiro P, Momennejad I.2021Interaction-grounded learning. arXiv.
[101] Zintgraf L, Devlin S, Ciosek K, Whiteson S, Hofmann K.2021Deep interactive Bayesian reinforcement learning via meta-learning. arXiv.
[102] Grover A, Al-Shedivat M, Gupta JK, Burda Y, Edwards H. 2018Evaluating generalization in multiagent systems using agent-interaction graphs. In Proc. of the 17th International Conf. on Autonomous Agents and MultiAgent Systems, pp. 1944-1946. Richland, SC: International Foundation for Autonomous Agents and Multiagent Systems.
[103] Battaglia PWet al.. 2018Relational inductive biases, deep learning, and graph networks. arXiv, 1806.01261.
[104] Son JY, Bhandari A, FeldmanHall O. 2021Cognitive maps of social features enable flexible inference in social networks. Proc. Natl Acad. Sci. USA 118, e2021699118.
[105] Perra N, Fortunato S. 2008Spectral centrality measures in complex networks. Phys. Rev. E 78, 036107.
[106] Lancichinetti A, Kivelä M, Saramäki J, Fortunato S. 2010Characterizing the community structure of complex networks. PLoS ONE 5, e11976.
(参考文献可上下滑动查看)
未来智能实验室的主要工作包括:建立AI智能系统智商评测体系,开展世界人工智能智商评测;开展互联网(城市)大脑研究计划,构建互联网(城市)大脑技术和企业图谱,为提升企业,行业与城市的智能水平服务。每日推荐范围未来科技发展趋势的学习型文章。目前线上平台已收藏上千篇精华前沿科技文章和报告。
如果您对实验室的研究感兴趣,欢迎加入未来智能实验室线上平台。扫描以下二维码或点击本文左下角“阅读原文”
