清醒和非快速眼动睡眠EEG微状态序列的频率分析
摘要
大多数脑电(EEG)微状态分析都是在清醒状态下进行数据采集,而现有的睡眠研究主要集中在空间微状态特性的变化以及相邻时间点之间的微状态转换上,睡眠状态下脑电微状态研究尚且不足。本研究旨在对清醒和非快速眼动(NREM)睡眠阶段的非平滑EEG微状态序列进行更广泛的时间序列分析。对不同的时间尺度采用不同的分析方法,短时间尺度可以采用马尔可夫检验来评估,中间时间尺度可以用熵率来评估,长时间尺度可以用频谱分析来识别微状态的特征频率。研究发现从清醒期到N3期的振幅逐渐下降,微状态平均持续时间的增加是一个渐进的现象,可以用转移概率矩阵的弛豫时间来描述微状态动力学的连续减慢。有限熵率表明,随着警觉水平的降低,微状态序列的可预测性增强(随机性降低)。因此,在清醒状态下不存在马尔可夫性质,但在睡眠阶段N3中,10/19个被试的微状态序列符合二阶马尔可夫过程。通过比较微状态序列的时滞互信息系数与脑电信号的自相关函数来进行谱微状态分析。本研究发现在所有警觉状态中的周期性微状态行为与清醒时的α频率、N1的θ活动、N2的睡眠纺锤波频率以及N3的δ频段有关。总之,EEG微状态是一种具有振荡特性的动态现象,在睡眠中减慢,并与几个睡眠阶段的特定脑电频率相耦合。
前言
清醒和睡眠的脑电图(EEG)分类是基于EEG频率、特定的EEG波形(顶尖波,睡眠纺锤波,k-复合波),将警觉状态划分为清醒、非快速眼动(NREM)睡眠(N1-N3)和快速眼动(REM)睡眠(美国睡眠医学学会,2007年)。在这里,本研究分析了清醒状态和NREM睡眠中脑电微状态的时间序列特征,尤其是振荡特征。
脑电微状态是由表面EEG测量并通过聚类算法识别的瞬态、准稳定的电场形貌。微状态序列是EEG数据集的时间序列表征,在这些序列中,对数十到数百毫秒的连续block进行微状态标记。聚类原理是将具有高度空间相似性的脑电地形图进行分组,以获得相对于底层脑电数据集的最大可解释空间方差百分比。聚类的最佳簇数可以通过多种成本函数来确定。许多研究都发现了四个最佳簇,并且这些微状态图具有高度一致性。这种高度可复制的地形图通常用字母A-D来标记。这些地形图的极性遵循一般模式:左枕到右额(图A),右枕到左额(图B),枕区到额区(图C),以及一种极性介于图C和径向对称模式之间的地形图D。在不同的研究中,这四种地形图的全局解释方差(GEV)在58%到84%之间。
在过去的几十年里,微状态特性已在许多实验条件下得到了验证,尤其是对健康和多种神经精神疾病(如精神分裂症、早期精神病、自闭症、嗜睡症和不同形式的痴呆症)的研究。既往关于清醒状态下微状态的研究表明,微状态A和B类分别与认知和感觉加工有关,微状态C和D类分别与突显网络和注意网络有关。尽管这些关系仍然存在争议,但同时也是当前研究的主题。
睡眠状态下的微状态研究发现,困倦和睡眠期间也存在A-D这四种典型的微状态。Brodbeck等人(2012)对清醒和NREM睡眠阶段的全频段EEG(1-40Hz)进行了微状态分析,研究结果发现微状态A-C的地形图非常相似。N2和N3睡眠阶段的微状态图D呈现出更圆(径向对称)的模式。微状态持续时间随着睡眠阶段的加深而增加,同时伴随全局场功率峰值之间时间间隔的增加。这些发现都表明,睡眠期间的微状态动态普遍减慢。时间属性是从一个时间点到下一个时间点转换的微状态转移矩阵推导出来的。本研究使用时滞互信息分析描述了振荡微状态动力学,发现当考虑非平滑的原始微状态序列时,微状态在清醒状态下的发生频率是主导EEG频率的两倍。例如,具有主导10Hz α活动的EEG产生的微状态序列在微状态之间(间隔为50ms(20Hz)、100ms(10Hz)及其倍数)具有较大的互信息值。睡眠脑电图显示了困倦时的θ波节律、N2期的睡眠纺锤波、深度睡眠(N3)时的慢波δ节律等特征性振荡现象,但目前不清楚清醒状态下的振荡微状态动力学是否也发生在睡眠中。此外,熵率和振荡微状态动力学受认知任务的调节,这些指标反映了认知负荷和认知控制之间的平衡。然而,我们不知道这种解释是否与我们对睡眠期间认知能力下降的理解相一致。
本研究的目的是测试不同睡眠阶段下微状态序列的信息内容和周期模式。将微状态解释为大规模网络活动的电生理关联,本研究假设(i)在觉醒-睡眠过渡期间,觉醒相关的微状态频率会随着觉醒特定网络活动的减弱而消失,(ii)随着睡眠特定振荡现象的出现,睡眠相关频段中可能会出现微状态振荡。本研究的方法确定了微状态序列是否具有周期性,因此能够跟踪大规模网络活动的时间行为。将微状态序列的时滞互信息函数(AIF)作为自相关函数(ACF)的广义形式,并将其与脑电信号的自相关函数(ACF)进行比较。通过ACFs和功率谱密度之间的对应关系,可以将EEG频谱峰与微状态AIF进行比较。
材料和方法
数据
本研究分析了32名健康受试者(年龄范围:19-27岁,平均年龄:23岁,20名女性,12名男性)参与无任务同步EEG-fMRI研究的EEG记录。纳入至少达到N2睡眠阶段且警觉状态持续时间≥105s的受试者。其中共19名受试者进入N3期睡眠。清醒状态为闭眼的无任务状态。对于每个被试,不同警觉状态下的数据片段被调整为相同的长度。在所有受试者中,数据集的总时长在105-210s之间(平均:157.97s)。EEG预处理步骤包括校正MR梯度和心电伪迹,降采样至250Hz,并进行平均重参考。本研究使用6阶巴特沃斯滤波器进行0.5-20Hz的带通滤波。
睡眠评分
根据美国睡眠医学学会标准划分睡眠阶段。对睡眠纺锤波进行手动标记,N2期睡眠纺锤波对应的微状态序列也进行单独的分析。睡眠纺锤波段(n=32)的持续时间为544ms-1844ms(均值:951.4ms)。
频率分析
为了总结所有通道的EEG频率信息,对每个数据集进行主成分分析。仅保留第一个成分,并使用Welch方法(Hann窗,4096ms)计算其功率谱密度(PSD)。每个频段的相对功率由该频段所覆盖的PSD面积除以总PSD面积确定。PSD是自相关函数(ACF)在时域的频域等效。ACF测量时间点t的信号xt与时间点t+τ的信号之间的线性相关性。时滞τ的ACF系数r(τ)定义为期望值E(xtxt+τ)。频率为f的振荡信号与振荡长度Τ=1/f具有周期自相关关系。例如,一个纯10Hz振荡(理想的脑电α节律)在100ms的倍数处具有局部ACF最大值,在50ms的奇数倍处具有局部ACF最小值(50,150,…ms)。
微状态分析
对于每个多通道EEG数据集,计算每个时间点的空间标准差或全局场功率(GFP):

其中vi(t)是指N个电极上测得的平均参考电位。图1显示了警觉状态下三个典型EEG通道上的代表性EEG波形,以及相关的GFP时间序列(红色)。在GFP函数的每个局部极值处,假定全局电场形貌是稳定的。采用Python编程语言中改进的k-means算法实现。聚类质心是给定对象的K个微状态图。与我们之前对睡眠脑电数据集的分析一样,将聚类分为K=4个微状态类别。k-means算法运行5次,并为每个受试者和睡眠阶段选择解释大部分空间方差的结果。从个体受试者地形图中,通过Koenig等人(1999)描述的全排列法计算组图,使用20次独立运行并保留具有最大解释方差的结果。分别计算每个警觉状态的微状态图。
将组图反拟合到原始脑电数据集中,得到微状态序列。在每个时间点,使用最适合当前EEG地形图的微状态类标签i∈{A,B,C,D},并测量微状态图与EEG地形图之间的平方相关系数。为了保持微态序列的时间结构不变,在每个时间点进行反拟合。不应用进一步的时间平滑方法。

图1.清醒(W)和NREM睡眠阶段(N1-N3)的脑电微状态。
微状态统计
本研究评估了被试在每个警觉状态的如下参数:
-每秒GFP峰值(PPS):每秒局部GFP最大值的平均值[1/s]。
-平均微状态持续时间(MMD):一个微状态地图在切换到下一个地图之前保持稳定的平均时间[ms]。
-全局解释方差(GEV):GFP加权后的空间方差百分比[%]。
-转移概率矩阵:一个4x4随机矩阵T,单个时间步长从微状态标签i切换到标签j的条件概率Tij=P(Xt+1=j|Xt=i),其中i,j∈{a,B,C,D}。
-转移概率矩阵T的弛豫时间(Trelax):从扰动到弛豫的速度。作为谱间隙的倒数,谱间隙定义为T的两个最大特征值之差。
采用单因素方差分析评估不同警觉状态下的差异,然后进行事后Tukey检验(α=0.05)。
马尔可夫替代数据
微状态标签的分布用π表示,P(Xt=i)指微状态序列Xt在时间点t上具有标签i∈{A,B,C,D}的概率。微状态分布π和转移概率矩阵(Tij=P(Xt+1=j|Xt=i))允许构建具有纯马尔可夫结构的替代序列。为了检测EEG微状态序列与马尔可夫过程的偏差,本文基于替代数据的零假设分布对经验微状态序列特性进行了检验。对于每个警觉状态,从所有被试的经验微状态序列中取T和π的平均值。在显著性水平α=0.05下,从n=500的马尔可夫替代值计算置信区间。
熵相关量
为了表针微状态序列中包含的信息量(或随机性),并从信息论的意义和独立于标签分配的方式描述它们的自相关结构,本研究分析了两个与熵相关的量,(i)有限熵率和(ii)自信息函数(AIF)。
这两个量都基于微状态标签分布。微状态序列可以用Shannon熵来表示:
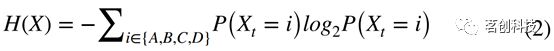
如果序列是由单一的、永不改变的微状态组成,则为最小值(H(X)=0);如果每个微状态分配到所有样本的1/4,则为最大值(H(X)=log(4))。Shannon熵是一种静态指标,因为它不包含任何关于样本时间顺序的信息。本研究使用有限熵率,利用条件熵引入时间依赖性。

该公式表示下一个微状态标签Xt+1的不确定性(或熵),根据前k个微状态的信息。计算涉及多元分布的Shannon熵(联合熵),本研究选择k=6个样本(24ms),根据既往研究,当序列长度k>6时,熵率并不可靠。具有较大熵率的序列Xt,其去相关时间较短。换句话说,如果产生的微状态标签具有较高水平的随机性和较低的可预测性,那么熵率将很高。另一方面,较低的熵率意味较高的可预测性。采用单因素方差分析评估不同警觉状态的熵率差异,并进行事后Tukey检验(α=0.05)。对于任意时滞t而言,时间点t和t+τ之间的两点相关性是通过微状态标签Xt和Xt+τ之间的共享(互)信息来测量的。互信息是两个随机变量之间线性相关系数的泛化,因此,时滞互信息系数的集合是时间自相关函数(ACF)的泛化,将其标记为自信息函数(AIF)。
![]()
AIF系数在时滞τ的大小表明时间t的微状态对时间t+τ的微状态的预测性能。在信息论语言中,它表示通过对Xt进行条件化后,Xt+τ的不确定性(随机性)减少了多少。AIF在规律间隔中的峰值表明微状态的周期发生率,就像ACF局部极值代表周期性脑电信号一样。
AIF峰值测试
为了检验AIF曲线的峰值(局部最大值),本研究构建了一个额外的替代检验。如果AIF曲线下的归一化面积(AUC)在时滞系数τ(即[τ-w,τ+w])附近的窄窗口内明显大于替代数据AIF下的相应面积,则AIF曲线在时滞系数τ处具有显著的峰值。面积相对于AIF曲线下的总面积进行归一化。
检验AIF a(t)在多个时滞系数
![]()
是否有显著峰值,其归一化常数为A=∫a(t)dt。使用Mann-Whitney U检验对该统计量的值与n=10时马尔可夫替代值的零分布进行比较。使用的经验窗半宽为w=8ms(2个样本)。本研究预计AIF峰值与ACF的局部极值一致。因此,对于警觉状态W、N1和N2,本研究测试了ACF的前两个局部极值给出的峰值位置τk。对于N3,只有第一个ACF最小值落在分析范围内τmax=1000ms,并用τk表示。例如,具有10Hz频谱峰值的EEG数据集产生的ACF的前两个局部极值位于50ms和100ms,因此使用τ1=50ms和τ2=100ms进行AIF峰值测试。
马尔可夫检验
该转移矩阵捕获了一阶马尔可夫过程描述的微状态序列特征。而马尔可夫性质描述了时间序列对过去的记忆程度。零阶马尔可夫过程可满足零假设,即当前状态Xt的信息不会影响到下一个状态Xt+1的转移概率。在一阶马尔可夫过程中,转移概率P(Xt+1|Xt)依赖于Xt,而不依赖于Xt−1或更早的样本。高阶马尔可夫过程的规则遵循相同的模式。如前所述,在每个警觉状态下测试了0、1和2阶的马尔可夫性质,显著性水平为α=0.05,并进行Bonferroni校正。所有方法都是用Python编程语言实现。
结果
微状态统计
不同警觉状态下的微状态如图1所示。通过Pearson相关系数测量不同状态下的相似性(表1)。在所有睡眠阶段,睡眠图和清醒图之间的平均相似度至少为80%。与清醒状态相比,图D在N1(62.2%)和N2(53.8%)中相似性最低,图A在N3(71.0%)中的相似性最低。图D的不同之处可以从视觉上描述为在睡眠中比清醒状态下更圆。表2进一步总结了微状态属性,以所有受试者的平均值和90%置信区间表示。单因素方差分析显示,所有参数的警觉状态存在显著差异。随着警觉性的降低,每秒GFP峰值数(PPS)减少,而微状态的平均持续时间增加。随后GFP峰值之间的时间间隔不断增加,表明脑电频率向较低的方向转移。
表1.睡眠N1-N3期与清醒期图形的相似性[%]。
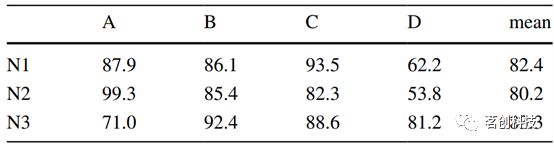
表2.微状态基本属性。

事后成对Tukey检验显示,所有警觉状态之间的PPS值有显著差异。对于MMD值,只有睡眠阶段N1和N2之间的微状态A、C和D之间的比较没有显著差异。GEV比较显示,图C解释了所有警觉状态下的最大方差百分比。总GEV值为58.9%(N1)~67.4%(N3)。
弛豫时间和熵率
单因素方差分析显示,在不同的警觉状态下微状态转移矩阵的弛豫时间存在显著差异。平均弛豫时间随着睡眠深度的增加而增加(W:4.5;N1:5.7;N2:6.4;N3:10.8)。熵率的变化趋势则相反,从清醒状态到深度睡眠状态呈下降趋势(W:0.86;N1:0.74;N2:0.69;N3:0.48位/采样间隔)。对于这两个参数,警觉状态之间的所有两两比较均显著(事后Tukey检验)。MMD、转移矩阵弛豫时间和熵率之间的关系如图2所示。为了更好的可视化,本研究绘制了与弛豫时间相反的谱间隙。对MMD值进行半对数缩放后,变量熵率和谱间隙几乎随MMD的增加而线性下降,呈现出近似对数的关系。谱间隙和熵率值以相同的速率下降。警觉状态由不同颜色和形状分开表示。

图2.微状态序列随着警觉性的降低而减慢。
马尔可夫性质检验
对清醒休息阶段和睡眠阶段N1-N3的微状态序列进行了0、1和2阶的马尔可夫性质检验。所有受试者在四种警觉状态下均拒绝了0阶和1阶马尔可夫过程的零假设。所有受试者在清醒和睡眠N1和N2阶段均拒绝二阶马尔可夫性质的零假设。在睡眠阶段N3中,9/19例被拒绝。因此,阶段N3中的10/19(52.6%)个微状态序列可描述为二阶马尔可夫链。对于独立的睡眠纺锤波,所有受试者(32/32例)均拒绝零阶马尔可夫性,1/32例受试者拒绝一阶马尔可夫性,任何一例受试者均无法拒绝二阶马尔可夫性(0/32)。
微状态频率分析
本研究测试了微状态序列是否包含与脑电频谱相关的特定频率。对于每个警觉状态,本研究比较了脑电功率谱密度(PSD)、脑电自相关函数(ACF)和相应的微状态自信息函数(AIF)。并计算多通道脑电信号数据集的第一主成分PSD和ACF曲线,将所有通道汇总为一维信号。将ACF曲线叠加到微状态AIF上,直接比较局部最大值和最小值。为了比较微状态序列与等效一阶马尔可夫过程中存储的信息量,本研究增加了从马尔可夫替代数据计算得到的95%置信区间的AIFs。
清醒状态
清醒阶段的分析见图3。PSD(左)在9.5Hz处有一个频率峰值,在正常成人α频率范围内。相对α频段(8-13Hz)功率占总功率(0.5-20Hz)的41%。对应ACF的第一个局部最大值(右侧黑色曲线)位于100ms处,第一个局部最小值位于52ms处。这两个ACF局部极值与微状态AIF的前两个局部极值重合(红色曲线)。AIF的前5个峰位于马尔可夫替代数据定义的置信区间之外(蓝色区域),表明微状态序列在这些时滞包含的共享信息数量明显大于等效马尔可夫模型。本研究还应用了AIF峰值测试。该测试针对前两个AIF峰,并将它们与平均替代AIF(蓝色曲线)进行比较。它还表明,与一阶马尔可夫模型相比,周期性是显著的(p<0.001)。
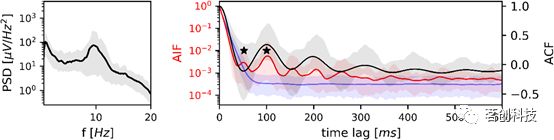
图3.清醒状态下微状态序列的PSD、ACF、AIF。
睡眠阶段N1
睡眠阶段N1的分析结果如图4所示。组平均PSD(图4a,左)呈近似单调衰减,无明显的频率峰。因此,组平均ACF和AIF均未显示出明显的周期性(图4a,右)。在所有n=32个被试中,本研究发现了一个在N1期间有显著θ振荡的被试。图4b展示了该分析结果,可以看到在6.3Hz处有一个明显的PSD频谱峰。ACF(黑色曲线)在76ms和148ms处出现了前两个局部极值。对于相同的时间滞后,微状态AIF(红色曲线)在马尔可夫替代定义的置信区间(蓝色区域)外具有局部最大值。AIF峰值测试相对于马尔可夫替代AIF(蓝色曲线)具有显著性(p=0.043)。
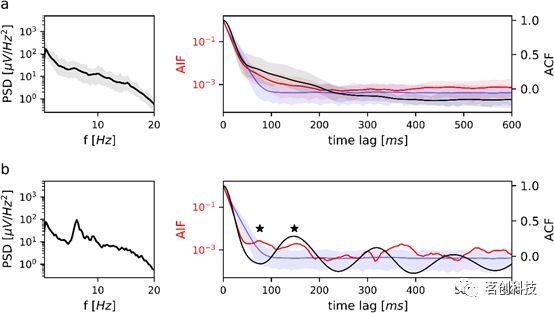
图4.N1期睡眠微状态序列的PSD、ACF和AIF。
睡眠阶段N2
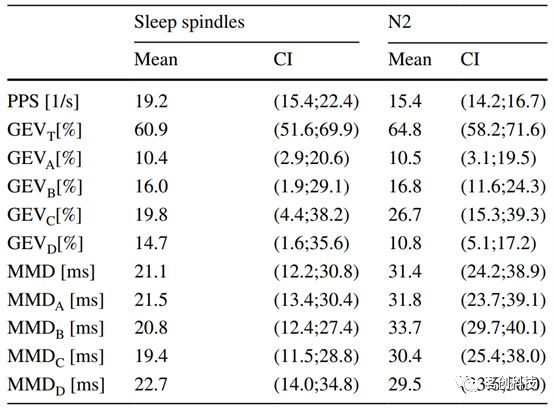
N2期的分析结果如图5所示。N2的组平均PSD(图5a)在δ、θ和α频段上单调衰减。在β低频段12.2Hz处观察到一个小峰,这是一个典型的睡眠纺锤波频率。图5a右侧的ACF(黑色)和AIF(红色)曲线没有明显的振荡。然而,在12.2Hz纺锤波频率(82ms)对应的时滞处,微状态AIF曲线高于马尔可夫置信区间(图5a中的箭头)。为了测试微状态是否随纺锤波频率变化,本研究分析了无睡眠纺锤波的片段,结果如图5b和表3所示。N2微状态图解释了60.9%的睡眠纺锤波段方差(GEVT)。跨微状态类的单因素方差分析显示GEV值之间存在显著差异(p=0.004),而事后Tukey检验显示只有图A和图C存在显著差异。此外,本研究发现每秒平均出现19.2个GFP峰(PPS),微状态平均持续时间为21.1ms。分离的睡眠纺锤波在12.5Hz处有一个频谱峰(图5b,左),在40ms和72ms处出现一个ACF局部极值。AIF峰同时出现滞后,但其绝对值没有超过马尔可夫替代数据(蓝色区域)的信息量。虽然睡眠纺锤波微状态序列(红色)对应的AIF位于马尔可夫替代置信区间(蓝色阴影区域)内,但其形状与平均替代AIF(蓝色曲线)明显不同。本研究使用AIF峰检验对红色曲线中的表观峰进行统计显著性检验,发现时间滞后40ms和72ms时的峰具有显著性(p<0.001)。这表明睡眠纺锤波微状态序列的AIF峰是实证脑电数据所独有的。在马尔可夫替代数据中从未观察到类似的峰值。

图5.N2期睡眠微状态序列的PSD、ACF和AIF。
表3.睡眠纺锤波分析。
睡眠阶段N3
N3期睡眠微状态序列的PSD、ACF和AIF如图6所示。左:半对数坐标下的平均功率谱密度(n=19)。右:脑电自相关函数(黑色)和微状态自信息函数(红色)。马尔可夫替代数据以95%置信区间(蓝色阴影区域)和平均替代AIF(蓝线)表示。PSD以高δ功率为主,在10.3Hz处有一个较小的峰值。ACF的第一个局部最小值是440ms(1.1Hz)。AIF峰值位于468ms(1.1Hz),相对于马尔可夫置信区间具有显著性,而替代AIF呈单调衰减;AIF峰值测试对AIF峰有显著性意义(用星号标记),并在下图显示了PSD、ACF、AIF及其置信区间(5-95%)。
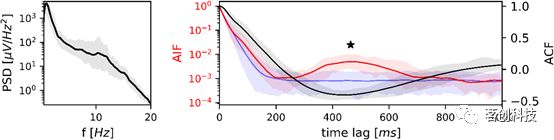
图6.睡眠阶段N3的频率分析。
结论
脑电微状态是一种动态的、周期性的现象,其频率变化与不同警觉状态下的脑电主导频率密切相关。本研究证明了微态动力学在整个非快速眼动(NREM)睡眠阶段持续减慢,并且当传感器水平的EEG信号显示出明确的频谱峰时,就可以观察到与α、θ、δ和睡眠纺锤波频率相关的振荡微状态动力学。这些发现表明,在单个电极上观察到的振荡可以扩展到大规模脑功能网络的振荡活动。
参考文献:Wiemers, M.C., Laufs, H. & von Wegner, F. Frequency Analysis of EEG Microstate Sequences in Wakefulness and NREM Sleep. Brain Topogr (2023). https://doi.org/10.1007/s10548-023-00971-y