title: 文献笔记-Where, When, and How Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasersinfection
date: 2019-11-22 12:00:00
type: "tags"
tags:
- 天然免疫
categories: - 文献笔记
注:以前看综述的时候,我喜欢把原文逐字翻译为汉语,效率比较低,现在我就采用笔记的形式的来看综述,按照原文的结构,把关键点记录下来,再加上自己的理解即可。
文献信息
Hailing Shi, Jiangbo Wei, Chuan He,Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers,Molecular Cell,Volume 74, Issue 4,2019,Pages 640-650,
ISSN 1097-2765,https://doi.org/10.1016/j.molcel.2019.04.025.
摘要
细胞RNA会天然地经历各种化学修饰。这些经化学修饰的核苷酸又赋予了其自身结构的多样性,从而在各种有序的代谢与机体的功能方面发挥着各种调控功能,进而影响了基因的表达。随着对回贴(mapping)位点修饰的全转录组测序方法的发展,对精确修饰进行检测和定量的高灵敏质谱方法的进步,以及参与修饰的“效应器”(effectors,包括各种能改变各种修饰位点的酶,例如写入器(writers)和擦除器(erasers)和能识别化学修饰位点的读取器(readers))理解的加深,最近几年有关RNA修饰的研究呈现暴发式地增长。然而由于RNA种类庞杂,表达水平有差异,因此这给RNA修饰的研究带来了很大的挑战,另外,RNA的修饰还与细胞类型,组织特异性,以及修饰效应器的定位有关,这又增加了研究难度。我们在这篇综述里,重点阐述了最近几年关于-甲基化转换酶(m6A, )功能的研究进展,强调了环境(context)对RNA修饰调控和功能的重要性。
前言
早期研究发现,m6A主要发生在(G/A)(m6A)C序列上。最近的研究发现,m6A主偏向于出现在终止密码子和3'UTR上,因此我们可以推断,m6A可以影响基因的表达。
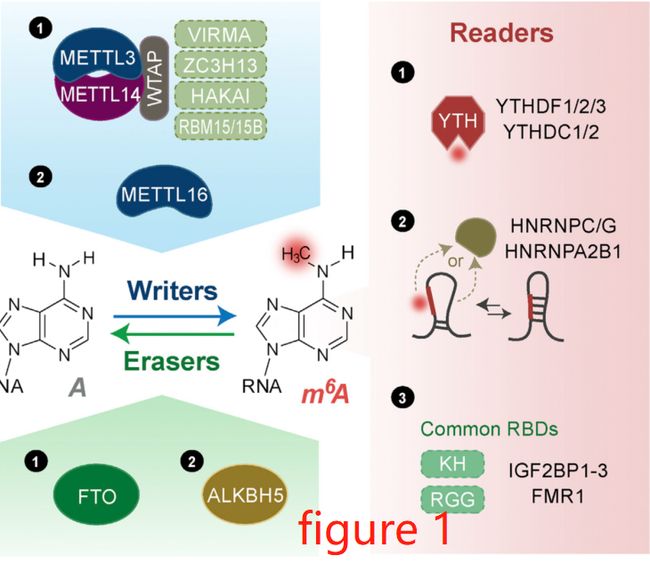
m6A途径的效应器(effectors)包括写入器(writers),擦除器(erasers)和读取器(readers),其中写入器往核苷酸上添加上甲基,擦除器反之,即去掉核苷酸上的甲基,读取器则是能够识别那些核苷酸上含有甲基的序列,它们的成员如下图的Figure 1所示:
机体的整个生物学环境会影响m6A通路,并且决定了m6A的功能。
一方面,不同的细胞,以及环境刺激能影响m6A这些
效应器自身的表达水平,PTM以及细胞定位。例如,在多数细胞系中,m6A的去甲基化酶(demethylase)FTO主要位于细胞核,并且参与5%-10%的的mRNA的m6A去甲基化,但是在某些淋巴瘤细胞系中,这个比例达到40%。
另一方面,RNA分子自身的异质性又增加了m6A的研究难度,这是因为:
①RNA种类繁多,包括mRNA,tRNA,rRNA,以及其它大量的非编码RNA;
②不同类型的细胞中的RNA丰度不同;
③RNA存在着复杂的二级结构;
④RNA存在着不同的转录状态(例如非活跃时,RNA与组蛋白紧密结合,转录活跃时,则与组蛋白松开);
⑤RNA中的顺式/反式作用元件(例如一些RNA结合蛋白,RBP, RNA binding proteins)会调控m6A效应子与RNA底物。因此说,m6A的活动与环境紧密相关。
m6A甲基化的写入器
m6A是通过一个复合物加到mRNA上的(Figure1),这个复合物有多个亚基,包括METTL3,METTL14,WTAP,VIRMA,ZC3H13,HAKAI,RBM15/15B。其中包括以下成员:
- METTL3/14,全称是methyltransferase-like 3/14,即甲基转移酶3/14, 这两个甲基转移酶是这个复合物的核心成员,其中MELLT3起催化作用,MELLT14促进复合物与RNA结合。
- WTAP,全称是Wilms tumor 1-associating protein,其功能是结合METTL3/14,诱导它们向底物招募以及定位。
- VIRMA,全称是Vir like m6A methyltransferase associated,功能是将m6A与3'UTR结合。
- ZC3H13,全称是zinc finger CCCH-type containing 13,功能是诱导写入器复合物向核内转移。
- RBM15/15B全称是RNA binding motif protein 15/15B,功能是与尿嘧啶富集区域结合,促进某些RNAs的甲基化。
METTL3的功能
METTL3在各类细胞中的分布不同,例如在HeLa细胞中,METTL3/METTL14会形成二聚体结构与WTAP产生相互作用,然后进入细胞核形成斑点(speckle)。METTL3和WTAP中含有NLS(nuclear localization signals),NLS的突变会导致复合物无法入核。METTL3还存在于一些细胞质中,例如在一些癌细胞系,包括HeLa,MDA-MB-231,MOLM13细胞系等。还有一些情况,例如在小鼠皮质神经元中,METTL14位于细胞质和细胞核。现在还不清楚为什么METTL3的定位为什么会出现如此差异。METTL3/METTL14的比例在不同的细胞系中也有差异,这可能会影响METTL3的定位。
写入器的招募导致了转录本上的m6A的特异性,这一招募过程可能是由TF或表观遗传标记介导的,如下下图的Figure 2所示:
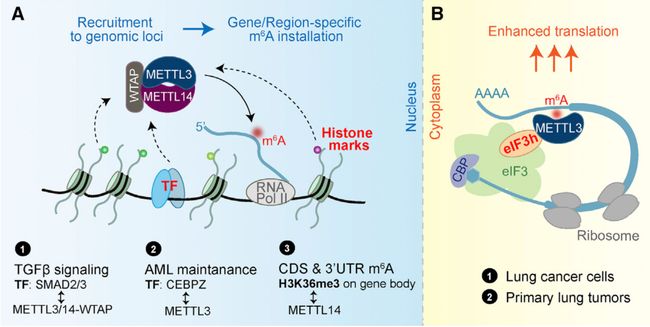
METTL3在细胞核中的作用
当出现热休克时,METTL3就会定位到染色质的热休克相关的基因上,m6A就会被添加到热休克转录本上,从而诱导这些转录本在应激压力下被清除。UV诱导DNA破坏时,METTL3/14就会在2分钟内定位到UV诱导的损伤位点,同时伴随着m6A活动的增强。在人类多能干细胞的TGF-信号转录通路转导方面,活化的SMAD2/3会与METTL3/14-WTAP相互作用,诱导m6A添加到特定的转录本上。在AML细胞中,一部分METTL3会通过METTL14非依赖方式与CCAAT增强子结合蛋白zeta(CEBPZ)的启动子区域结合。在HepG2细胞中,H3K36me3能通过与METTL14相互作用招募写入器,诱导mRNA的CDS和3'UTR上添加m6A。
METTL3在细胞质中的促进翻译作用
METTL3还能作为一个潜在的m6A读取器,在Figure 2B中,我们可以看到,在肺癌细胞中,METTL3此时并没有发挥催化作用,而是在细胞质中锚定到了3'UTR上,促进了一个报告mRNA的翻译。进一步的研究表明,METTL3是通过eIF3h相互作用来促进翻译的。
METTL3蛋白自身会通过PTM或与其它蛋白质相互作用来发挥调控功能,例如人类的METTL14能够诱导METTL3的丝氨酸399磷酸化。人类的METTL3会出现SUMOylation修饰,导致METTL3/14的活性降低。但这些现象的机制还不清楚。
序列和结构特异性诱导的甲基化
METTL3/14复合物偏爱RRACH模序(R=A或G;H=A,C或U),METTL16则偏爱另外的序列与结构。METTL16被验证的两个特异性序列是U6(U6 small nuclear RNA, snRNA)和人类MAT2A(methionine adeno-syltransferase 2A)上的一个发夹结构(hp1),MAT2A编码S-腺苷甲硫氨酸合成酶(S-adeno-sylmethionine synthetase, SAM synthetase)。有实验分析发现,METTL16偏爱一个形成茎环结构膨大处的UAC(m6A)GAGAA序列。EMTTL16介导的MAT2A甲基化是SAM稳态的负反馈信号。最近发现了ZCCH4是一个新的m6A读取器。
m6A读取器
m6A读取器通过作用于含有m6A的mRNA来发挥作用,Figure 1中可以把读取器分为三类。
第一类读取器含有YTH结构域(YTH的全是YT521-B homology),这些成员包括YTHDF1-3(YTH domain family 1-3),YTHDC1-2(YTH domain containing 1-2)(参考Figure 1)。细胞质中的YTHDF2会通过招募CCR4-NOT腺嘌呤酶复合物来诱导靶转录本的部分降解。细胞质中的YTHDF1和YTHDF3能促进HeLa细胞中靶转录本的翻译。细胞核中的YTHDC1有多个作用,包括招募某些剪接因子调控mRNA的剪接,促进mRNA的输出,加速某些转录本的降解。YTHDC2调控mRNA的稳定,翻译以及精子形成。
第二类读取器是HNRNPs,全称是heterogeneous nuclear ribonucleoproteins,成员包括HNRNPC,HNRNPG与HNRNPA2B1,它们能调控靶转录本的剪接,加工,它们能与RNA的某些结构结合,这些结构是由m6A介导形成的,因为m6A会重构局部的RNA结构,调控RNA-蛋白质之间的相互作用,这一现象称为m6A开关(m6A-switch)。
第三类读取器含有相同的RNA结合结构域(RBD, RNA binding domains),例如KH结构域(K homology),RNA识别模序(RNA recognition motif),以及RGG( arginine/glycine-rich)结构域,它们都能与含有m6A的mRNA结合,包括FMR1和IGF2BP1-3。FMR1(Fragile X mental retardation 1)含有3个KH结构域,1个RGG结构域,它有可能通过与YTHDF1和YTHDF2相互作用来影响RNA的转移,RNA的稳定。IGF2BP1-3(insulin-like growth factor 2 mRNA-binding proteins 1–3)能通过m6A的方式稳定靶转录本(YTHDF2与之相反,它是降解靶转录本)。IGF2BP蛋白功能的核心是KH3-4结构域,这是与m6A结合的关键区域。
Prrc2a是最近发现的一个读取器,它与髓鞘形成有关,具体机制不明。
为什么读取器会结合一些m6A位点
以下为作者的几个猜测。
第一,读取器有可能与其它的RBP相互作用,从而被招募到mRNA的不同区域。IGF2BP1-3和YTHDF2通过在不同位点来调控mRNA的稳定性,例如IGF2BP1-3结合到3'UTR,而YTHDF2结合到CDS区。而RBP对RNA的识别取决于多个因素,例如结合位点的序列,序列的侧翼以及RNA的二级结构。因此RBP与读取器蛋白质的相互作用有可能涉及了m6A位点的特异性。
第二,m6A位点的密度和序列也可能与之有关。
第三,读取器蛋白会在细胞区室的特定位点富集,因此会偏向与局部一些RNA类型结合。例如,YTHDF1-3,FMR1和HNRNPA2B1位于细胞的应激颗粒核心地区。在乳腺癌细胞中,YTHDF1-3,HNRNPK和IGF2BP2-3位于细胞的突起(protrusion)。
甲基化可以促进翻译或影响某一转录本的稳定性,这取决于揎细胞环境下哪个读取器占据主导地位,如Figure 3所示:
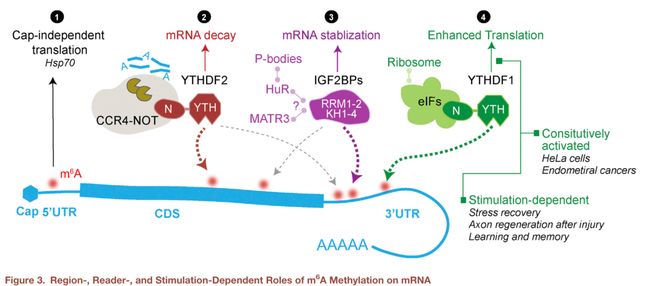
读取器对外界刺激的应答
在某些刺激下,例如热休克应激或病毒感染会诱导细胞质中的YTHDF进入细胞核。在热休克出现的几小时内,YTHDF2的转录本与蛋白水平都上升,并且多数YTHDF2转移到细胞核内。有人认为,YTHDF2核心的YTHDF2会与去甲基化酶FTO竞争,阻止那些热休克应答基因5‘UTR的去甲基化,从而增强这些基因在细胞质中非加帽方式的翻译(cap-independent translation)。体外实验发现,YTHDF2能抑制FTO的去甲基化活性。在Vero细胞感染实验中,细胞感染了enterovirus type 71后,YTHDF1和YTHDF2会上调,并且在感染12小时后分布在细胞质,感染24小时后分布在细胞核,这一现象机制不明。
在有丝分裂后期的细胞中,当需要功能蛋白时,YTHDF1的翻译促进作用就变得特别明显(Figure 3)。在损伤诱导的轴突再生的背根节(DRG)模型中,再生相关基因大量地被甲基化,在这个恢复过程中,YTHDF1是蛋白质强烈合成所必需的条件;相比之下,YTHDF1在损伤前的基础翻译增强方面,作用很小。这一现象与最初在HeLa细胞中研究YTHDF1时类似,YTHDF1会促进一些转录本的翻译。在关于记忆研究方面,有研究发现,YTHDF1的缺陷会导致海马依赖性神经元功能出现障碍,恢复YTHDF1的蛋白水平,则能修复这种问题。YTHDF1依赖的翻译增强会在一些癌细胞中持续出现,而在其它一些细胞中则是由刺激诱导的,这一过程是如何触发的,还不清楚。
读取器的不同功能
YTHDF1-3的结构类似,它们都含有N末端低复杂序列,和C末端的保守YTH结构域。但是它们的功能并不相同,在不同的细胞中,它们有的能促进蛋白翻译,有的能抑制蛋白翻译,具体的案例可以参考原文。并且在不同的m6A阅读器蛋白之间还存在着交叉作用或竞争作用,即使在FTO和阅读器之间,也有着类似的作用。因为这些蛋白本身就构成了一个有相互作用的网络结构,因此要理解它们的环境调控作用对于相关的研究有着重要意义。
m6A擦除器
m6A甲基化可以被FTO或ALKBH5逆转,这两个蛋白也就是m6A擦除器。FTO是第一个被发现的m6A擦除器。FTO能够去甲基化m6Am,以及部分mRNA上的cap-m6Am,如Figure 4所示:

CLIP-seq验证了很多FTO的结合位点,这些结合位点包括tRNAs,snRNA等。FTO的空间分布也能发挥调控作用,FTO的N末端有一个NLS,它能部分地分布在细胞核中,也能分布在细胞质中,FTO的分布在不同的细胞系中有所不同,例如在AML细胞中的分布和HEK,HeLa细胞中的分布就不同,这可能与肿瘤的发生有关,FTO在AML细胞中的这种分布也有可能是环境依赖的,因为2HG(2-hydroxy-glutarate)能抑制FTO。
CLIP-seq的测序数据分析表明,在某些细胞系中,FTO偏爱结合GAC-和/或GGAC模序。有研究发现,FTO的表达和定位可能被PTM调控(例如Lys-216)。
位于mRNA的5'cap中的+1核糖上,它几乎是与内部m6A(internal m6A,这个内部指的是mRNA中间的部分)同时发现的。m6A可能占据了整个腺苷酸的0.1%-0.4%,它主要发生在第二个核苷酸2’-O-methylated()的帽子结构附近。只有不到30%的含有m6Am。m6Am的比例占到整个mRNA的m6A十分之一或更低(在一些细胞系中还要低)。m6A丰度要远高于,因此,虽然FTO更偏爱结合,但是FTO的主要靶点还是m6A。由于几乎所有的mRNA帽子结构都在胃镜核中被加帽蛋白结合,因此在细胞核中,cap-不太可能被去甲基化,这也反映了FTO对的局限,如Figure 4所示。现在研究发现的FTO还功能涉及:热休克应答,UV损伤应答,HCV感染等方面。
mRNA的甲基化不影响转录本的稳定
最初研究认为FTO介导的的去甲基化在功能上类似于DCP-2介导的去帽(decapping)化诱导mRNA的降解以及miRNA介导的mRNA降解。当敲低FTO后,FTO的靶mRNA出现了很多变化,这些效应被认为是由于的去甲基化导致的,但是FTO敲低与那些只含有m6A或转录本水平之间的相关却不支持这个结论,并且发现只有内部m6A修饰的转录本的变化与FTO的敲低相关。
后来发现了PCIF1是mRNA的甲基转移酶,它只加工,却不加工内部的m6A(internal m6A)。还有研究发现,PCIF1添加的并不改变基因表达的水平或转录本的稳定性,这也不支持刚开始的观点,即FTO介导的的去甲基化导致的mRNA降解。另外,有研究发现,当FTO敲低时,被认为还能促进那些含有mRNA的翻译效率,但也有一些研究发现了相反的现象,这一过程的调控也有可能是与环境有关。
ALKBH5的作用与底物
ALKBH5是第二个被发现的m6A去甲基化酶。ALKBH5在小鼠的精子形成中发挥了作用,ALKBH5和FTO的表达在不同组织中各异。例如,在小鼠中,Alkhb5主要表达在睾丸,而Fto主要表达在大脑,这可能与它们参与的不同生物学途径有关。在人源细胞系中敲低ALKBH5后,会诱导其靶RNA从细胞核转移到细胞质。几乎所有报道的ALKBH5功能研究都揭示了类似的分子途径,ALKBH5介导某些转录本的3'UTR m6A,其中就包括促进缺氧诱导的HIF依赖的乳腺癌干细胞表型,通过ALKBH5-FOXM1途径调节的胶质母细胞瘤增殖和肿瘤发生,调控雄性生殖细胞中长3'UTR mRNAs的剪接和稳定。
结论与展望
作者在结论部分中讨论了两个问题,第一个是m6A的调控,第二个是m6A未解决的一些问题。
其中m6A的调控功能可能受几个层面的影响,包括以下几个方面
第一,m6A的功能取决于细胞分化和细胞的发育状态。
第二,m6A的功能可能被环境因素或细胞信号触发。
第三,m6A效应子的定位也影响了m6A的功能。
第四,即使是同一个转录本,m6A精确的定位和其它的修饰也会影响其功能。
关于m6A的一些关键问题还没搞清楚,包括以下几个方面:
第一,m6A效应子针对转录本的选择性和m6A位点的选择性是如何实现的?
第二,m6A效应子(包括写入器,擦除器,读取器)是如何整合到不同的生物信号转导与调控过程的?