论文笔记:气候变化、水循环和植被动态
植被与水文,水循环
植被影响植物蒸腾,影响蒸散发,影响降水,影响径流和水储量,进而影响水循环,水资源

植被动态和水循环的交互:
目标:
- 未来气候条件下,植被如何调节水通量?
- 在气候引起的水胁迫下,植被如何变化?

水分传输:水力过程-水的潜在梯度驱动
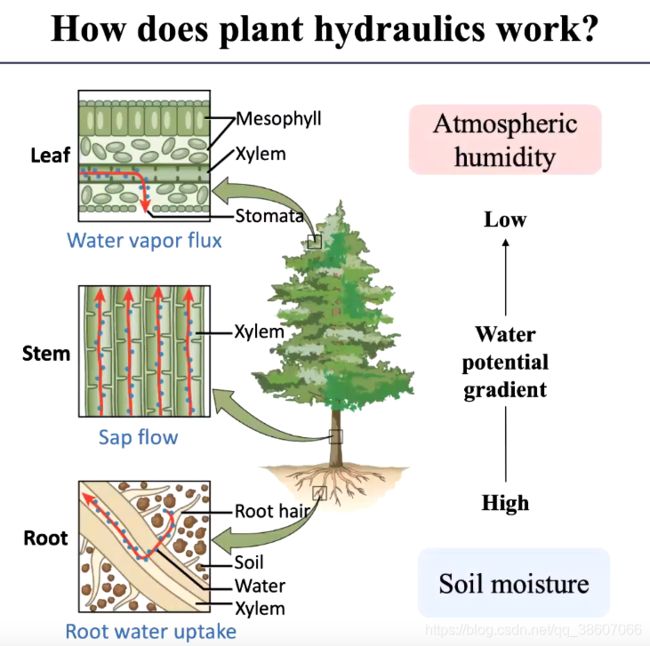
干旱对水分传输的影响:
干旱导致土壤水分下降,栓塞效应,导管输送水分的能力下降,

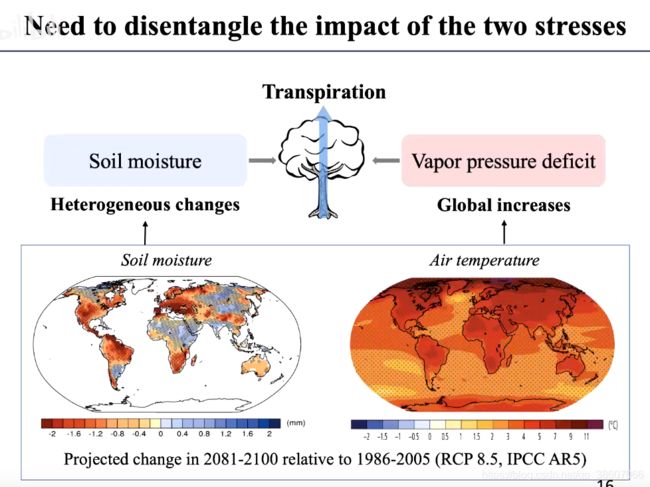
目前,土壤湿度和大气湿度VPD是高度相关的。
但在未来气候情境下,土壤湿度有升有降,但大气干旱普遍上升(气温上升)。
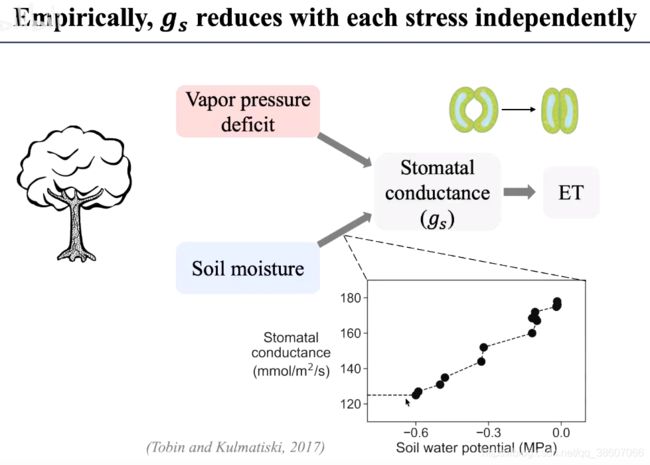

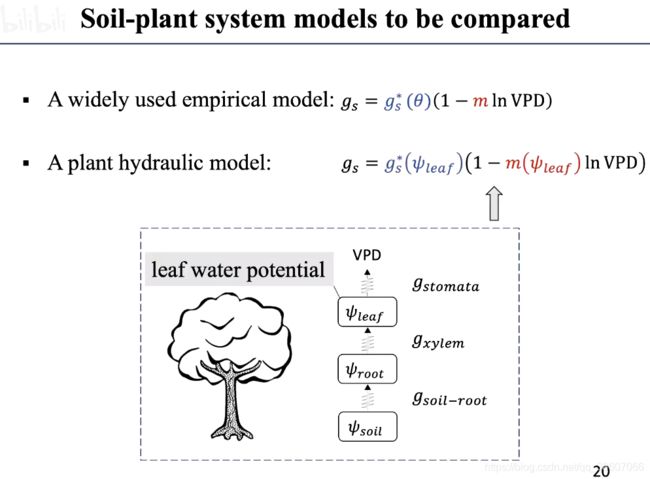
如何优化参数?


植被影响气候
植被通过对地表能量收支的影响来影响气候
反照率:被地面反射的太阳短波辐射的比例
反照率决定了被地面吸收的太阳辐射量,这些吸收的能量接下来作为长波辐射和潜热、感热的湍流通量输送到大气中去。
水体(湖泊、海洋)的反照率通常较低,吸收了大量的太阳辐射。
冰、雪的反照率一般较高,吸收的太阳辐射很少,以维持其存在的寒冷条件。
植被的反照率处于中等水平,枯叶草地、落叶林、暗针叶林的反照率依次递减。
当前的土地利用变化极大地改变了区域的反照率,通过增加裸露地表的面积。
土壤的反照率决定于土壤类型和湿度,但在干燥气候下通常比植被的反照率高。
因此,过度放牧通常增加反照率,降低地表吸收的太阳辐射和向大气输送的能量。这会导致冷却和下沉,因此潮湿的海洋空气不会被海风吹向内陆。这会降低降水和植被从过度放牧中恢复的能力。
许多陆面对气候的巨大反馈表明,陆面变化是区域气候变化的重要因素。
生态系统结构影响潜热和感热湍流通量输送到大气中的效率。
风从高的、不平坦的冠层吹过会引起机械湍流,从而增加了从地表到大气的热传递效率。
相比之下,光滑的表面往往会升温,因为它们只通过对流而不是机械湍流来传输热量。
植被结构对水、能交换效率的影响影响着区域气候。
亚马逊盆地25-40%的降水来源于陆地的蒸散发。
模型模拟表明,如果亚马逊盆地的森林全部转化为草地,这将导致该地的气候永久性的变暖变干。(森林变草地,蒸腾作用会减弱,潜热输送会减少,地表温度会上升。热带森林有助于保持地表凉爽和潮湿。)
草地的根系比树木较浅,吸收的土壤水较少,导致较低的蒸腾速率。
因此,草地会以显热的形式释放更多的吸收的太阳辐射,这将直接温暖大气。
仍存在不确定性,例如,云的变化对辐射强迫的影响有正有负,取决于云层的性质和高度。

由植被变化造成的反照率变化会生产放大效应的反馈。
例如,在高纬地区,在雪融化之前,树木覆盖的地表比冰雪覆盖的苔原吸收更多的太阳辐射。
模型的模拟结果表明,6000年前的林线北移,降低了区域反照率,增加的辐射吸收足以解释当时气候变暖的一半。
温暖的区域气候反过来有利于植被的再生长和林线的建立,提供了一个对区域变暖的放大的正反馈。
因此,预测未来气候对植被的影响也应考虑生态系统对气候的反馈。
反照率、潜热和感热通量的能量分配和地表结构也会影响向大气发射的长波辐射能量。
长波辐射取决于地表温度,当地表吸收了大量的入射辐射(低反照率),更少的水分去蒸发,或者有光滑的表面(热传递效率低——传递给大气的潜热和感热的湍流通量效率低),地表温度倾向于升高。
例如,沙漠的净长波辐射能量亏损很大,是因为其干燥、光滑的地表引起了高的地表温度,以及较少的水分蒸发以降低地表温度。
第四章:水和能量平衡
被太阳能所驱动的水循环是驱动其它生物地球化学循环的一个关键循环。本章描述了生态系统能量平衡和其它水循环调控机理
引言
水和太阳能对生命至关重要
水循环和能量循环紧密相连,不宜分开研究。
太阳能通过蒸散发将水从地表垂直输送到大气,驱动水循环。
蒸散发(evapotranspiration)=地表蒸发(evaporation)+植物蒸腾(transpiration)
蒸散发占到了湍流能量输送(潜热+感热)的80%以上,是地球能量收支的关键过程。
水循环通过溶解营养物质和在生态系统之间和内部输送来控制地球的生物地球化学循环。
一个焦点议题
水汽反馈:水蒸气是主要的温室气体,对太阳短波辐射透明,而吸收地球长波辐射,形成温室效应。
CO2和其它温室气体排放引起的气候变暖会增加大气中的水蒸气数量和提升大气捕捉长波辐射的效率。
这种水汽反馈解释了为什么气候对其它温室气体的排放如此敏感?
海平面上升:
主因是海洋的热膨胀,其次是冰川和冰盖的融化。
表面能量平衡
辐射收支
一个表面所吸收的辐射能量是入射和出射辐射的平衡
辐射收支:短波辐射,源于太阳
长波辐射,源于地球(所有物体)
净辐射Rnet

短波辐射的输入=直接辐射+散射辐射+反射辐射
shortwave input = direct radiation + diffuse radiation + reflected radiation
直接辐射:太阳辐射直接穿过大气层到达地表的部分(晴天的占比可达90%)
散射辐射:被大气中的粒子和气体所散射的短波辐射部分(阴天、污染重、太阳高度角小的黎明或黄昏比重大)
反射辐射:云层和周围景观单元所反射的部分
被吸收的入射短波辐射的比例取决于生态系统表面的反照率或短波辐射反射率。
例如:新雪的反照率很高(白、亮),湿土、湖面、海洋、针叶林树冠等反照率很低(黑、暗)。
反照率不仅取决于单个叶片、茎和土壤,也取决于生态系统的结构。
一个复杂的冠层的反照率低于单个叶片,一个叶片反射或传输的光会被另一个叶片吸收,作为一个整体,冠层捕捉光的效率更高。
例如:针叶林的冠层深且不平,其反照率低;作物、草地的冠层平滑,其反照率高。
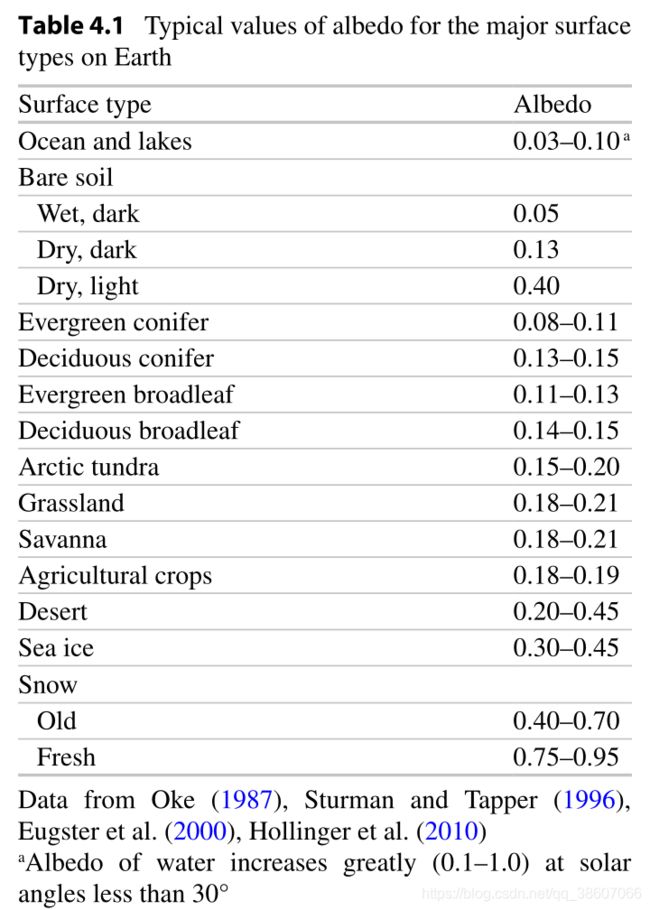
生态系统反照率的变化部分解释了为什么高纬地区比低纬地区暖化更快(极地放大效应):
气候变暖,冰雪融化,高反照率的冰雪地表被低反照率的土壤、森林代替,陆面吸收更多的辐射能量,地表温度上升,传输到大气中的能量也会增加。会形成区域性的气候变暖的一种放大的正反馈。
这一过程就是冰雪反照率反馈。snow (or ice) albedo feedback。
反照率随太阳辐射输入的短期变化而变化。
一天之内,树冠在正午比黎明或黄昏吸收更大比例的入射辐射(反照率更低)。
阴天比晴天吸收更多的散射辐射。
物体发射的长波辐射取决于其表面温度和发射率。
植被生态系统的发射率约为0.98,大多数吸收的辐射都会被发射出去,因此长波辐射平衡主要取决于天空的温度(决定了长波辐射的输入)和生态系统表面的温度(决定了长波辐射的输出)。

生态系统的长波辐射的输出决定于表面温度。
表面温度决定于表面接收的辐射数量+通过湍流过程传输给大气和土壤的能量
干燥表面和低蒸腾速率的叶片往往特别温暖,因为不会被水的蒸发冷却。
总之,短波辐射输入、反照率、表面粗糙度、表面温度是影响辐射平衡和净辐射的关键因子。
吸收辐射的分配
净辐射,生态系统吸收的辐射能量,大致通过非辐射过程从生态系统中输出出来的能量所平衡。
非辐射过程包括:
地表热通量G:传到能量给土壤
从陆表到大气的湍流输送:潜热LE(水的蒸散)+感热H(热的蒸散)=蒸散
少量(小于10%)存储在生态系统中的能量ΔS=光合作用产生的化学能+植物生物体的温度升高
存储在生态系统中的能量通过呼吸和生物体温度下降的形式释放出去。
尽管由光合作用捕捉的能量是驱动生态系统碳循环的主要能量动力,但它只占到整个生态系统能量收支的一小部分(小于5%)。
净辐射=潜热+感热+地表热通量+生态系统中存储的能量

地表热通量G:在多数生态系统,一天之内的地表热通量基本忽略不计,因为白天向下传递给土壤的热量与晚上向上传递给地表的热量相平衡。
地表热通量的量级决定于表层土壤和深层土壤的热梯度和土壤的导热系数。
最陡的热梯度和最大的土壤热通量出现在多年冻土区域。
例如:在北极,夏季约有10-20%的被吸收的能量用于融化冻土;但在冬季,当土壤重新冻结,这部分能量又返回到大气中。
湖泊和海洋在夏季也会存在地表热通量:太阳输入穿透到水面以下,水的高热容和湍流混合,高效地将水面的热量移走。
热量从地表向大气传递主要靠湍流。
湍流:在表层和自由大气(bulk air)之间的不规则速度的空气移动。
自由大气:不受到冠层强烈影响的冠层上方的空气。
产生湍流的过程有两种:
对流湍流convective turbulence:从表面到近表面空气1-2毫米以上的地方,由感热传到(扩散)引起。
温暖的空气膨胀,导致浮力增加,造成低密度的温暖空气上升,产生对流湍流。
机械湍流mechanical turbulence:更加高效地能量传递方式。当空气从不均匀的表面移动,其水平移动速度会不均匀的减慢。
空气动力学的粗糙 aerodynamically rough
从不均匀的地形和植被表面经过的气流产生了一个向下扫入冠层的空气旋涡,将自由空气输入进冠层,将冠层内的空气输送出去。这些涡流将能量从冠层表面传送走,并混合到大气中。
光滑的冠层比粗糙的冠层产生的湍流更少,热量传递效率更低。因此其表面温度会更高。
湍流不仅传递显热,也传递潜热。
表面的水分(湿度)决定了生态系统的感热和潜热的交互方式。
蒸腾吸热,降低表面温度,降低表面和大气之间的温度差,抑制感热通量。
感热通量温暖表面空气,提高空气的饱和水汽压,导致潮湿空气远离蒸发表面的对流运动。
潜热和感热都会提高水汽梯度,用以驱动蒸发。
表面湿度对bowen ratio影响很大。
bowen ratio:感热通量与潜热通量的比值-决定了生态系统能量收支和水收支的联系紧密程度。
bowen ratio越低,表明生态系统中蒸散占湍流能量交换的比重大,水收支和能量收支的联系更紧密。
bowen ratio在不同的生态系统之间差别很大:
- 湿生态系统以潜热为主,bowen ratio小于0.5。干生态系统以感热为主,bowen ratio大于0.5。
- 物种特征:生长旺盛、光合速率高的生态系统bowen ratio小,以潜热为主。例如落叶林的bowen ratio小于针叶林。
- 强风和粗糙的冠层:产生大气湍流,降低表面温度,从而降低感热通量
景观上生态系统的空间配置会影响能量分配,因为临近生态系统之间的热量对比会产生对流湍流。
生态系统边界处的湍流,以及随之而来的潜热和感热通量大于生态系统内部(中心)。
当在反照率和能量分配上具有强烈差异的生态系统斑块的直径大于10千米(行星边界层高度)时,这样的生态系统能改变中尺度大气环流、云和降水模式。
蒸散是生态系统中水和能量收支中最大的项之一,因此控制蒸散量级的因子决定了水循环和能量循环联系的紧密程度。
水的比热较大,加热1克水1℃所需的能量是加热同等质量空气1℃所需能量的4倍。
比热(specific heat):加热1克物质1摄氏度所需的能量。
水的相态变化会吸收或释放大量的能量。在20℃汽化1克水需要2.45MJ kg-1的能量,是把1克水加热1℃所需能量的580倍。
在蒸发表面和蒸腾叶片上,蒸散具有很强的冷却作用。与之相反,水汽凝结成云也会对大气有很强的温暖作用。
水汽压是大气中水分子施加的分压。
距蒸发表面非常近的空气在蒸发表面温度下接近饱和。
VPD(vapor pressure deficit)是相同温度条件下,实际水汽压和饱和水气压之差
VPD是蒸散的驱动力,实际上也是水分从土壤通过植物运往大气的驱动力。
生态系统水收支的综述
可用于支持生态系统生产力的水取决于输入和输出之间的平衡

生态系统就像是一个水桶,被降水填充,通过蒸散和径流排空。
蓝水:可以别社会利用的河流、湖泊、水库和底下含水层中的液态水。
绿水:土壤表面的蒸发+植物蒸腾
陆地生物圈绿水通量(60%)大于蓝水通量(40%)。
生态系统的水输入
对大多数生态系统而言,降水是主要的水输入
生态系统水输入的数量和季节性:降雪与降水不同
冠层截留:在多雾的生态系统中增加生态系统的水输入。但在少雾地区,冠层截留通常会减少由于冠层蒸发而进入到生态系统的降水比例。
生态系统内部的水运动
水运动从冠层到土壤
在密闭冠层的森林中,进入的降水有很大的比例落在冠层上。
冠层的水量平衡:落在冠层上的降水=直接蒸发返还给大气+叶片吸收+throughfall(透冠降水)+茎流stemflow(水从树茎到地面)
冠层截留:未到达地面的降水的部分(在浓密的森林,约战10-20%)。不等于落在冠层上的降水
不同生态系统,冠层截留和存储降水的能力不同,主要取决于冠层表面积,尤其是叶片表面积。
例如:针叶林通常比落叶林存储更多的水。
冠层截留的影响因素:林龄和附生植物负载,通过影响冠层表面积影响冠层截留。
茎流:
影响因素:树皮纹理,树干结构,中继线
草地的降水少于森林,但30-40%的降水被冠层截留。
总之,冠层截留减少了土壤水分输入。只有在有雾的情况下除外,冠层截留增加土壤水分输入。
土壤中水的储存与运动
土壤水分主要存储于土壤颗粒表面的薄水膜中
土壤持水能力取决于土壤总孔隙体积和周围颗粒的表面积。
孔隙体积又取决于土壤深度和孔隙所占土壤体积的比例。
土壤水分从高势能到低势能沿着梯度运动
水的能量状态取决于水的浓度和各种压力。
自然生态系统中的压力可用静水压力和基质作用力描述。
静水压力主要有:重力压力,它取决于高度+由蒸发和生物体的生理过程产生的压力。
基质作用力来源于细胞表面和土壤颗粒对水分的吸收。水膜越薄,水分子收到表面基质作用力的吸附就越强,越紧密。
水势water potential:水相对于土壤表面纯水的势能
总水势=pressure potential压力势能 +osmotic potential渗透势能+matric potential基质势能
压力势能:由重力和生物体的生理过程产生。
渗透势:反映了溶解在水中的物质的存在。
基质势:由表面吸收水分产生。
按照惯例,在无压力情况下,土壤表面纯水的水势为0.
如果比参考值大,水势为正;如果比参考值小,水势为负
在生态系统的多数部分,水势为负值。
与重力和基质力相关的压力梯度控制大部分水分在土壤中的运动。
土壤中的水流速度取决于驱动力(水势梯度)和水流阻力。
水流阻力取决于土壤的导水率和水流经的路径长度。
由于土壤质地和团聚体结构的不同,土壤倒水率有很大的不同。
雨水向土壤中的渗透不仅取决于导水性,还取决于土壤中由植物根系和土壤动物形成的裂缝或沟渠所形成的大孔隙的优先流preferential flow。
受重力影响,土壤中的水分向下流动,直到基质作用力大于重力,水分被吸收至土壤颗粒表面。
未被基质作用力截留的水分通过土壤排除,进入地下水,然后进入溪流和湖泊。
土壤的田间持水量是土壤重力水排出以后保留的水量。
田间持水量下,土壤的水势约为-0.03MPa,接近纯水的水势0MPa。
随着土壤变干,土壤水膜会变薄,剩下的水与土壤颗粒表面保持的更加紧密。
永久萎蔫点是指土壤变干,水势约为-1.5MPa,多数中生植物因不能从土壤中获取水分而枯萎。
旱生植物却能适应土壤干旱,当土壤水势低至-3到-8MPa之间,依然能够从土壤中吸收水分。
土壤变干,水膜变薄的第二个结果是干旱土壤中的水不能直接穿过充满空气的土壤孔隙,不得不沿着更长、更曲折的路径穿过土壤孔隙边缘的水膜。
因此,随着土壤变干,土壤的水力传导性迅速下降。
持水能力water-holding capacity=田间持水量-永久萎蔫点=植被可利用的水plant-available water
从土壤到根系的水运动
水分从土壤到蒸腾植物根部的移动,通过从高水势到低水势流动的方式。
只要根系的水势低于周围土壤,水就会从土壤向根系运动。
水沿着水势梯度向根系运行,导致了临近根系的土壤颗粒水膜变薄。剩下的水被土壤颗粒表面更加紧密的吸附,水势变小。
根系附近土壤水势的局部降低,导致水沿着土壤薄膜想植物根系运动。
水存储的变化
当水分输入超过水分输出时,多余的输入水可以用来补充土壤水分及地下水
人们通过改变植被以及其扎根深度,或直接抽取地下水补充日常用水等方式改变了底下水库
径流
径流量相当于降水输入、储水变化与蒸散失水的差值
径流对降水变异的反应要比蒸散作用强烈得多
基本径流base flow:缓慢流动的地下水为小河提供了持续的水源基本径流,即便是在没有降水的时期也一样
植被对径流量的影响非常强烈
植被通过影响蒸散来影响水循环,从而影响径流
流域的水量平衡:降水=蒸散+径流+水储量变化