20世纪70年代中期B淋巴细胞杂交瘤技术问世,由此制备出的单克隆抗体是抗单个抗原决定簇的抗体,具有高度的特异性和均一性,且能大量制备,从而为抗体的研究和应用带来了突破。以抗体为载体耦联放射性同位素毒素和药物的导向药物或“生物导弹”风靡全球。
经过近十余年的研究和临床试验,人们发现这些“导弹”药物离真正的应用还有一段距离。抗体的临床应用结果反映出单克隆抗体药物存在的一些问题,主要有:
①鼠源性抗体用于人体后产生人抗鼠抗体(HAMA反应);
②由于鼠单抗是异源蛋白,在体内很快被清除;
③由于抗体相对分子质量较大抗体分子到达靶部位的量不足;
④抗体本身的效应功能不强等,使抗体治疗的效果不理想。
但随着人们对抗体基因结构与功能的深入了解和分子生物学技术的发展及向各学科渗透,应用DNA重组和蛋白质工程技术可以按人们的意愿在基因水平上对抗体分子进行切割、拼接或修饰,重新组装成新型抗体分子,并可赋予抗体分子新的生物学活性。因此基因工程抗体比天然抗体具有更广阔的应用前景。
南京铭研生物提供重组抗体制备服务
服务优势:
● 一站式服务:抗体测序,亲和力测定,亲和力成熟,人源化,抗体表达及稳定系构建等一站式抗体相关服务
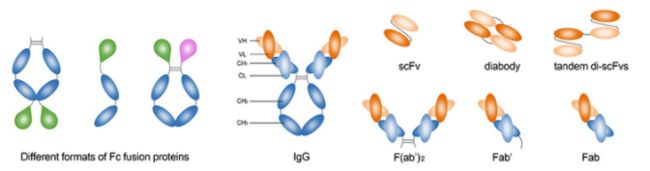
● 我们可以生产多种形式的抗体,包括全长抗体,scFv,Fab,sdAb,isotype/subtype及多种形式的Fc融合蛋白。
欢迎咨询官网:www.merrybio.com.cn
鼠抗体人源化及人源抗体制备技术
针对鼠抗体用于人体后产生HAMA反应,影响靶向性和疗效,以及异源蛋白在人体内被很快清除的问题。利用分子生物学手段和技术进行鼠抗体人源化。
目前人源化的方法有多种,最早的人-鼠嵌合抗体就是将鼠抗体可变区基因与人抗体恒定区基因相连后得到的融合蛋白。它完整地保留了亲本鼠单抗的特异性和亲和力,同时大大降低了鼠单抗的异源性。
已被美国FDA批准的抗CD20抗体 Rieux-imab( IDEC-O2B8)就是一个人-鼠嵌合抗体。将鼠抗体高变区的3个CDR移植到人抗体的相应部位,得到的人源化抗体称为重构抗体或改型抗体,其人源化程度超过90%。但要保留亲本抗体的特性有一定困难,因为抗体与抗原结合时,虽然可变区的CDR起很大作用,但抗体骨架区的结构也会通过与CDR相互作用影响分子结构,因此成功的例子不多。
近年来计算机模建技术的发展和蛋白质晶体结构数据的不断增加,使表面氨基酸残基“人源化”技术有可能用于抗体的人源化改造不同种属间免疫球蛋白的同源性极高,尽管它们的轻链属于不同型别,其CDR的长度也不一致,但暴露于表面的氨基酸残基的位置和数量却很保守。这些表面残基的组成模式在种系内相当保守,而种系之间互不相同,这种差异是免疫原性的来源。将鼠轻、重链可变区组成的Fv段表面暴露的骨架区氨基酸残基中与同者改为人抗体中的氨基酸,使Fv的表面残基人源化,消除其免疫原性而Fv的整体空间构象,从而保留其抗原结合部位的结构。表面残基人源化后抗保留了亲本抗体的特异性和亲和力降低了免疫原性。
20世纪90年代初,人们采用PCR方法克隆全套抗体重链和轻链可变因,并将它们重组到原核载体上,通过大肠杆菌直接表达有功能的抗体分子再用抗原筛选与之结合的抗体克隆。这种抗体库技术使抗体的制备技术发的飞跃。继而人们又利用噬菌体表面展示技术将抗体基因与丝状噬菌体外壳基因融合,使抗体片段表达于噬菌体表面,构建成噬菌体抗体库用抗原筛选达相应抗体的噬菌体克隆,经扩增可获得大量抗体。抗体库技术的明显优点是:
①扩大了筛选容量,用杂交瘤技术一般筛选能力在上千个克隆以内,而抗体选106以上克隆,有利于获得一些稀有抗体;
②抗体库技术可直接克隆到抗因,避免了杂交瘤分泌抗体不稳定而丢失的危险;
③所得抗体基因可进一步进造,以增加特异性和亲和力。
利用噬菌体抗体库或合成抗体库技术人们可以获得人源抗体。但由于不能完全解决经免疫的人B细胞来源所得抗体往往力较低。
在抗体库技术基础上,利用表位印模选择( epitope imprinted select)也称作抗原导向选择技术,可以将鼠抗体转变为人抗体。该方法是将小鼠抗链可变区或轻链可变区基因与人抗体轻链可变区基因文库或重链可变区基因混合成为“鼠-人”抗体库。用抗原进行筛选,挑出有结合活性的克隆,该克隆鼠抗体的重链基因与人抗体轻链基因组成或小鼠抗体轻链基因与人抗体重链组成。再将由此得到的人轻链或重链可变区基因与另一条人抗体链(重链或轻链)可变区基因文库混合,构成人抗体库。再次用抗原进行筛选,所得与抗原结合体克隆由人轻链、重链可变区组成,由此可得到特异性和亲和力与亲本鼠单抗相同的人单抗。
随着基因敲除技术的成熟,人们可以将小鼠免疫球蛋白基因组敲除( knock-out),将人免疫球蛋白基因组植入( knock-in)。这样的人源化小鼠用抗原免疫后产生特异性人源抗体,而人抗体可直接从免疫动物的血清中纯化,或获得足够数量对抗原特异的B细胞,用于制备单克隆抗体或进一步用噬菌体抗体库技术制备出基因工程抗体。
改变抗体分子大小增强靶向性
抗体分子大小、与抗原结合价数直接影响抗体的穿透性、廓清率及在血池和靶部位的比例。用分子生物学手段可以获得完整抗体和各种抗体片段。
有一定应用价值的抗体片段,例如,一个轻链可变区和一个重链可变区相连的单链抗体scFv、两个单链抗体相连的 diabody、两个单链抗体通过类似铰链区的接头与恒定区CH3相连的minibody。
一般情况下,抗体分子越小越容易通过血管壁,但在血液中被清除的速度也快。完整抗体穿透性差,但在血液中的半衰期长,可达几十个小时。diabody和 minibody则介于单链抗体和完整抗体之间。以抗体在靶部位与血液中的比值(T/B)而言, diabody和 minibody明显高于完整抗体和单链抗体。T/B比值的增高意味着抗体在靶部位浓度高粑向性好。此外,由于diabody和nobody都是二价的抗体,它们与抗原的结合能力远高于单链抗体,目前认为是较理想的免疫显像或治疗用抗体形式
增强抗体效应功能
由于天然抗体主要是通过调理作用、ADC或依赖补体的细胞毒效应起到杀伤靶细胞的作用。因此,天然抗体的细胞毒效应有限。下列几种途径可以增加抗体对粑细胞的杀伤。
免疫结合物( Immunoconjugate)
免疫结合物是以抗体或抗体片段为载体连接放射性核素、药物或毒素构成这些细胞毒性物质大大增强了抗体杀伤靶细胞的能力,抗体与放射性核素结合物的主要优点是不需要内化,并且由于射线作用有一定范围因此可以克服抗原的异质性,而且可以根据需要选择相应的核素,进行免疫显像或治疗。抗体药物结合物也不需要内化,有些可以产生旁效应,而且效应谱和毒性谱较清楚。抗体一毒合物也称免疫毒素,常用的毒素有白喉毒素、麻毒素、相思豆毒素和假单孢杆外毒素,它们的作用机制清楚,细胞毒效应强。上述免疫结合物中有些已被FDA批准成为药物。
免疫细胞因子( immunocytokine)
免疫细胞因子是将抗体与细胞因子基因连接后表达的融合蛋白,抗体的特异性与细胞因子的有效免疫刺激活性有机结合起来。免疫细胞因子没有常规化疗的不良反应。因为它们的作用原理与化疗药物不同,后者主要是杀伤分裂细胞,因此分裂快的正常细胞也受到损伤,而免疫细胞因子能识别肿宿细胞上的特异性抗因此避免了对其他组织细胞的伤害。通过细胞因子对免疫系统的激活,可以控制肿瘤转移和杀灭残留病灶。
双特异性抗体
双特异性抗体又称双功能抗体,是同一抗体的两个抗原结合部位分别针对两个不同的抗原,在结构上是双价的,而与抗原结合的功能是单价的。
双特异性抗体可以用化学交联、细胞融合和基因工程等方法获得,由于它可以同时与两种抗原发生反应,并使之交联,因而可介导标记物与靶抗原的结合,或使某种效应分子定位于靶细胞,此外,又由于它与抗原结合的单价性,不易引起靶抗原的调变,从而可提高抗体的某些生物学效应。双特异性抗体重链的可选择性使其Fc片段与Fc受体结合的能力明显减弱减少了该抗体在体内的非特异性分布,双特异性抗体的这些特性使它在诊断和治疗上有广泛的应用前景。
目前作为治疗肿瘤用的双功能抗体常采用抗肿瘤相关抗原(TAA)及抗CD3或抗TAA及抗CD16。这类双特异性抗体在荷瘤动物模型中无论是抑瘤试验还是杀伤试验均获得良好结果。无论采用何种免疫活性细胞的效应分子,其杀伤均无MHC限制。这为临床应用提供了许多方便。目前已有一些双功能抗体在进行临床试验。
细胞内抗体( intrabody)
一般的抗体在细胞内合成后分泌到胞外,如果在抗体基因的N端或C端加入引导序列就能使抗体表达定位在亚细胞部位,如胞质线粒体、内质网或细胞核部位。这种在细胞内合成并作用于细胞内组分的抗体称为细胞内抗体或内抗体。细胞内抗体可以提供一种其他方法不能做到的研究分子功能的方法。它可以在细胞内抑制病毒复制和抑制生长因子受体或癌蛋白表达,因此有作为基因治疗的应用前景。研究较多的是用细胞内抗体抑制人类免疫缺陷病毒1型(HV-1)和抗肿瘤。
抗体作为治疗制剂最早用于病原微生物感染引起的疾病,现在已发展到抗肿瘤、抗移植排斥抗血栓形成及自身免疫性疾病的治疗等方面。理论上讲,任何一种抗原都可以刺激机体产生相应的抗体如果该抗原分子参与机体的生命活动或疾病的发病过程,则相应抗体可通过中和拮抗、封闭、阻断等生物学效应起治疗作从目前抗体临床应用的情况来看,抗体作为治疗制剂用量较大因此提高抗体表达量降低生产成本等都是亟待解决的问题。此外,能否用小分子模拟抗体的作用也是值得研究的方向。