遗忘的生物学机制
遗忘和记忆的习得和巩固一样,是大脑管理记忆的生物系统的一部分。
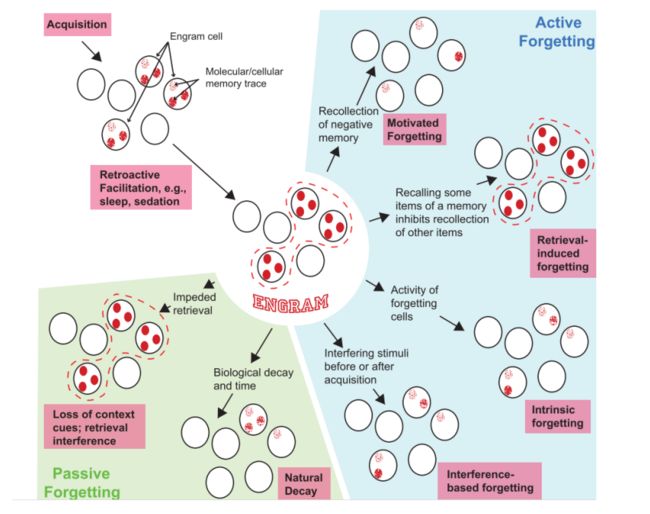
图1
(包括了记忆的获得保存以及主动和被动的遗忘。Engram细胞从外界获得记忆,通过睡眠等方式追溯加强。左下为被动遗忘;右半部分为主动遗忘的机制)
图1说明了记忆的形成和engram神经细胞的工作机制。在通过学习活动记忆后,engram神经细胞内活动发生变化,以此来保护细胞不受刺激,使记忆痕迹保持。被动遗忘有2种机制:因线索阻碍而导致时间线索的丢失和其他类似的记忆的干扰;另一种则是随时间推移导致的“自然衰减”。
而主动的遗忘——按图1分为在回忆中的消极记忆会导致遗忘或对其他记忆的抑制;由遗忘相关细胞导致的内在性遗忘;以及基于干扰的遗忘。当认知机制自愿参与削弱记忆痕迹时,就会发生有动机的遗忘。作者主要从此视角通过遗忘细胞的活动和大脑的内在生化和分子途径来介绍活动性的遗忘。
利用艾宾浩斯记忆曲线说明习得和巩固的强化力与遗忘在形成记忆图中的弱化力之间的平衡。遗忘很可能是发生在一小部分的记忆细胞内的。部分engram神经细胞和神经电路断开,使得记忆回路不完整,无法触发回忆的神经机制。
动机性遗忘(主动遗忘)是指在我们自己的认知控制下的遗忘过程,通常包含情感动机的遗忘过程(内疚,悲伤等)。自愿抑制记忆可能会破坏形成稳定的engram神经回路的过程,导致遗忘。
通过类别学习和回忆的实验可推测出检索引起的遗忘是由干扰产生的。实验表示该类遗忘可能有很多机制,但无论检索引起的遗忘的机制如何,随着时间的推移,记忆的自发恢复都可能会发生。作者假设大脑还具有利用诸如用于获取和巩固的信号系统来侵蚀记忆痕迹的固有生物能力,这类主动遗忘是通过过分子信号系统和大脑固有的神经元回路发挥作用的。
关于非遗忘(non-forgetting)的记忆痕迹。许多实验表明,记忆作为行为分析的指标,尽管会因为遗忘而遭受损坏,但可以通过微弱的“提醒”训练等来恢复。以此有观点认为尽管在没有提醒的情况下完全失去了行为表现,但engram回路或其部分组件在不同的时间内保持足够完整,就可以通过简短的提醒来重建;但同样有反对论点声称,在观察时被观察者本身有一个极其微弱的engram回路,记忆的恢复是由于通过提醒或其他方法加强了原本微弱的回路,并驱动了回忆机制。
另外,一些证据表明,生化记忆痕迹持续存在于记忆可以成功检索的时间之后。Li and Richardson做了大鼠幼鼠的tone-electric shock实验,并在实验前向大鼠NMDA受体注射拮抗剂来阻止tone-electric shock的记忆获得。但虽然幼崽已经忘记了行为分析的线索,记忆可以在之后的条件反射中重新获得。
还有非遗忘的极端观点设想, 所有曾经在大脑中表现出来的记忆片段都会继续被表现出来,而遗忘只是因为回忆不成功。成功的回忆可能取决于语境,情绪状态,能量状态,竞争心理活动的强度以及其他未知因素。
关于记忆痕迹的主动遗忘。之前的研究虽然干扰和其他机制会导致一些记忆失败,但无法解释所有的原因。一般来说,生物过程具有单独和专用的合成和降解途径,生物系统稳态的概念为内在遗忘过程的概念提供了额外的逻辑。从生物学的角度,内在的遗忘可被视为一种稳态机制,使大脑恢复到基本状态,从这个角度来看,由内在遗忘机制介导的遗忘可能是大脑的默认状态。
使用果蝇的开创性研究,已经确定了主动遗忘的几种分子和细胞机制。果蝇的研究表明了依赖Rac1的遗忘。在果蝇中,有证据表明Rac1至少介导了四种类型的嗅觉记忆主动遗忘,包括内在遗忘、干扰性遗忘、记忆遗忘和逆转学习激活遗忘。Rac1依赖的遗忘提供了一些经验:信号通路的激活可以用来获得记忆和主动遗忘;记忆形成和遗忘是独立的,遗忘不是通过逆转获取途径发生的(实验显示急性操纵Rac1对记忆形成没有影响, 但对遗忘有特殊作用);强的短期记忆不会自动合成长期记忆。
通过以果蝇为模型的研究,可以发现学习会激活Rac1蛋白来调控短期记忆的遗忘。并且已经有研究显示,脊椎动物(以小鼠做的实验)同样有着由Rac1调节的主动遗忘机制,表明主动遗忘可能在进化意义上有着高度的遗传保守性。
主动遗忘与脑部疾病之间的关系。虽然每种疾病都有一些独特的生理原理,但突触结构和功能的改变似乎是连接这些疾病的共同线索。作者猜想主动遗忘机制的改变是影响自闭症谱系障碍(ASD)的基础。基于小脑样本基因表达分析的一项系统生物学研究得出结论,Rac1在与ASD相关的神经病理事件中起着关键作用。有证据表明,人类受试者和小鼠遗传模型中的某些类型的ASD与逆转学习或主动遗忘缺陷有关。遗传缺失或基因敲除P-Re x1在小鼠海马的CA1区域产生ASD样行为,包括受损的社会互动、逆转水迷宫的学习缺陷和灭绝训练中抵抗恐惧的记忆调节。这些行为变化可能是由于活动性遗忘受损、记忆更新的失败。
遗忘细胞很可能是用于形成许多不同类型记忆的电路的一部分。我们接受细胞的存在,这些细胞形成记忆痕迹来储存在engram电路,同时还接受电路中的神经元,这些神经元将记忆形成的基本信息传递给这些engram细胞。作者推测,这些“学习电路”大多数内都有相关的遗忘细胞,来侵蚀在Engram细胞中形成的记忆痕迹。分析主动遗忘的生物学原理可以获得丰富的信息,并重新理解大脑如何管理它每天处理的大量信息。
2018年3月15日,清华大学生命科学学院和清华-IDG/麦戈文脑科学研究院钟毅教授研究组在Neuron杂志在线发表了题为Active Protection: Learning-Activated Raf/MAPK Activity Protects Labile Memory from Rac1-Independent Forgetting的研究论文,报道了学习/训练自身会通过激活Raf/MAPK信号通路来主动地保护所新形成的短期记忆,以阻止记忆由于意外干扰而快速遗忘。换句话说,不管是生理性的或通过药物激活这个机制,你刚记住的英语单词就不会因为突然来的电话或是一场火灾而忘掉。
早在1885年,德国心理学家艾宾浩斯(Hermann Ebbinghaus)就描绘出了人类大脑对于新习得事物的遗忘曲线,从此开始了人类探寻遗忘奥秘的旅程。在随后的一百多年间,许多学者利用实验心理学的方法,试图寻找遗忘的产生机制,也随之诞生了多种理论去解释遗忘现象,但最终都未能获得统一解释。遗忘现象作为人类记忆的主要特征,虽然在心理学中被广泛地研究报道,但是在神经生物学的研究中几乎无人问津,俨然成为了被“遗忘”的角落。主要原因可能是遗忘过程广泛被认为是记忆的反面,是一个被动的过程(记得差则忘得快,记得好则忘得慢),从而忽略了遗忘自身可能存在独特的分子机制。
直到2010年,钟毅教授课题组首次发现了主动遗忘机制,即学习过程本身会激活遗忘信号分子通路来主动地遗忘所形成的记忆。这种主动遗忘机制并不影响学习自身效果的好坏,而是特异影响已经学会的记忆的遗忘速度的快慢。这种主动遗忘的机制存在的意义很可能是为了及时抹除不需要的记忆,保证大脑留取足够的存储空间,从而不影响下一次的学习效果(Shuai et al., 2010; Davis and Zhong, 2017)。此项研究具体使用了果蝇为动物模型,发现学习会激活小G蛋白家族中的Rac1蛋白来调控短期记忆的遗忘。近些年的钟毅课题组的研究发现,果蝇中Rac1调控的主动遗忘机制也在哺乳动物小鼠模型中存在,表明主动遗忘功能在进化上存在高度的保守性(Liu et al., 2016)。随着研究的深入,钟毅课题组又进一步发现,像短期记忆一样,中长期记忆也存在主动遗忘机制。该机制是由和Rac1同一家族的Cdc42蛋白来负责调控的(Zhang et al., 2016)。这一研究发现也表明,遗忘很可能如同记忆现象一样具有高度的复杂性和精密性。
虽然Rac1介导的主动遗忘机制可以很好的解释为什么短期记忆会被很快速的遗忘,但是即使阻断Rac1介导的遗忘,研究者们还是发现短期记忆仍然会发生一定程度的遗忘。这个实验数据提示还有独立于Rac1遗忘机制存在的未知遗忘通路。在寻找其他遗忘机制的过程中,钟毅课题组发现了Raf介导的主动保护机制。首先,研究者发现学习过程会主动激活Raf/MAPK信号通路。延长这一信号通路激活时间后,研究者们观察到短期记忆也被显着的延长,而不影响记忆的形成过程。相反,缩短这一信号通路激活时间后,短期记忆会发生更快速的遗忘,而且同样不影响记忆的形成。这表明,Raf/MAPK通路会抑制一种遗忘机制。接下来,令人好奇的事情是,Raf抑制的遗忘和Rac1介导的遗忘是不是同一种机制呢?非常神奇的是,当研究者同时抑制Rac1遗忘机制和Raf抑制的遗忘机制后,果蝇的本来只可以维持数个小时的短期记忆在3个小时过去后都没有发生一丁点的遗忘,甚至在一天之后依然记得大量的短期记忆。这个研究事实提示,Raf/MAPK所抑制的遗忘可能是独立于Rac1通路存在的另外一种遗忘机制。这一猜测也得到了相应数据的支持。
然而,为什么动物需要同时拥有两套独立的机制来遗忘短期记忆呢?学习完成后,多种干扰会加速短期记忆的遗忘。常见的联想学习(associative learning)是指包括人类在内的动物可以通过将一个较为中性的条件刺激(conditioned stimulus)偶联一个强烈的非条件刺激(unconditioned stimulus)而学会两种刺激的相关性。例如,给小狗喂食(非条件刺激)时同时摇铃铛(条件刺激)。多次训练后,小狗可以学会铃铛声就代表着食物。学习后的干扰简单分为联想学习干扰(如相似学习干扰和逆转学习干扰)和非联想学习干扰(如电刺激、环境温度剧变和学习相关的嗅觉刺激)。研究者发现,调控Rac1只影响联想学习干扰造成的短期记忆遗忘,而不影响非联想学习干扰造成的短期记忆遗忘。而调控Raf/MAPK信号通路刚好相反。研究进一步发现,Raf/MAPK信号通路是通过激活其下游分子non-muscle myosin II (NMII)进而影响微丝骨架变化而维持突触结构的稳定性来发挥主动保护作用。
对于遗忘的探索过程为理解学习的作用提供了新的思路。学习过程除了获取记忆之外,它至少还触发了两件事情:一方面,学习过程会激活Rac1/Cdc42的活性来执行主动遗忘功能,这种“需要”的遗忘功能会帮助动物清理之前存在的不需要的记忆,为后续新记忆形成腾取空间;另一方面,学习会激活Raf/MAPK的活性来行使主动保护功能,加强记忆的稳定性,防止记忆意外丢失。这种主动保护机制会帮助动物免受“不需要”的干扰因素的破坏。因此,主动遗忘和主动保护的相互协调作用对于维持记忆的平衡和稳定性具有重要作用。
清华大学钟毅教授研究组致力于对遗忘功能和机制进行深入系统的研究并取得了一系列重要研究进展。2010年,Cell文章首次提出“主动遗忘”假说并找到短时程记忆的“遗忘”分子是Rac1; 2011年,PNAS文章报道了痕迹条件反射记忆的遗忘分子机制;2015年,PNAS文章报道了主动遗忘的神经环路机制;2016年,Cell Reports文章报道了Cdc42介导中长时程记忆的主动遗忘机制;Current Biology文章报道了Rac1介导的主动遗忘机制在小鼠中也是保守的;PNAS文章报道了多种自闭症风险基因的突变会通过抑制Rac1介导的主动遗忘功能而使得动物模型产生记忆灵活性失调的表型;2017年,Neuron展望文章概述了遗忘的生物学研究现状以及未来可能的研究方向;2018年,Neuron文章报道了独立于Rac1主动遗忘机制之外的其他遗忘机制会受到Raf/MAPK介导的主动保护机制的抑制。
作为世界性难题的阿尔兹海默病会使得患者发生严重的记忆能力损伤而影响正常的工作与生活。随着多种新开发药物的失败,我们究竟是否可以研发出改善记忆的药片也成为研究者们关心的焦点。过去此类药物的开发都把关注点放在改善记忆形成的方面,而独立于记忆形成的遗忘机制的发现为我们提供了新的思考角度。