转录组+代谢组助力大环内酯类抗生素对藻类抑制作用机制的研究
泰乐菌素(TYN)是一种常用的大环内酯类抗生素,可有效对抗革兰氏阳性菌,并可作为兽药用于预防肠道和呼吸道感染。TYN残留物可以通过在农田中施用粪肥和泻湖废水进入环境,值得注意的是,全球地表水中已检测到浓度高达3.06 μg L-1的TYN。已有研究表明,TYN对藻类的生长和发育产生不利影响。TYN能够抑制铜绿微囊藻和羊角月牙藻的叶绿素合成,进而抑制其光合作用。此外,TYN降低了铜绿微囊藻和羊角月牙藻的生长性能和增殖能力。
羊角月牙藻是经济合作与发展组织(OECD)推荐的生态毒理试验模式藻类,因为它分布全球,生长迅速,对有毒物质敏感。此外,已经完成了羊角月牙藻全基因组测序,并研究了暴露在抗生素中的分子机制。在本研究中,我们利用转录组和代谢组的综合分析来揭示TYN暴露下羊角月牙藻生长抑制的潜在分子机制。本研究发现,在TYN胁迫下,羊角月牙藻生长抑制与光合作用、DNA复制和能量代谢相关的分子通路失调有关。
技术路线
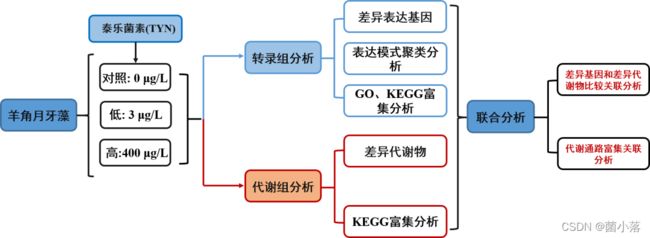
研究结果
- TYN对羊角月牙藻生长的影响
在本研究中,以3μg L-1(低处理组)和400μg L-1(高处理组)浓度的TYN处理羊角月牙藻7天(图1)。处理4天后,高TYN处理组的羊角月牙藻相比于对照组减少了67.8%。处理7天后,低处理组和高处理组的生长均受到显著抑制,相比于对照组分别减少了26.3%和58.3%。
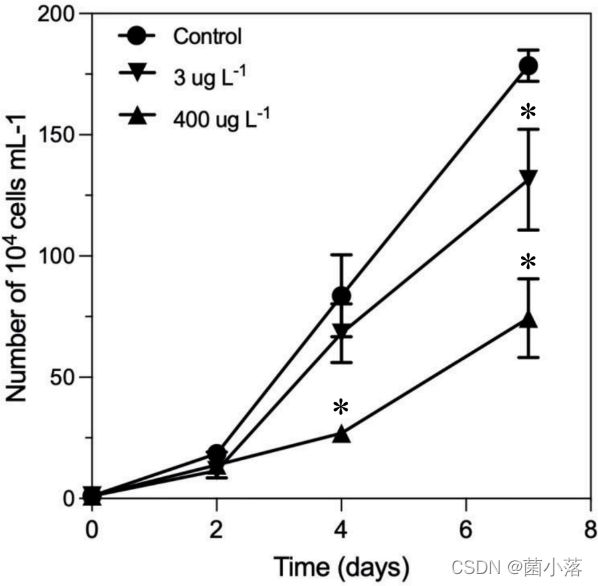
图1 不同浓度TYN处理7天后的羊角月牙藻生长曲线
2.差异表达基因
相关分析表明,各处理组重复间具有良好的一致性,各处理组3个重复中基因表达的相关系数均大于0.99(图2A)。值得注意的是,相同处理组的重复样本在PCA中有很好的聚类,其中PC1和PC2分别解释了93%和4%的变异(图2B)。与相关分析和PCA的结果一致,heatmap显示低TYN处理和对照的基因表达模式非常相似,但与高TYN处理的基因表达模式有明显的不同(图2C)。与对照组相比,低、高TYN处理组分别检测到上调差异表达基因329、2383个,下调差异表达基因248、3055个(图2D)。
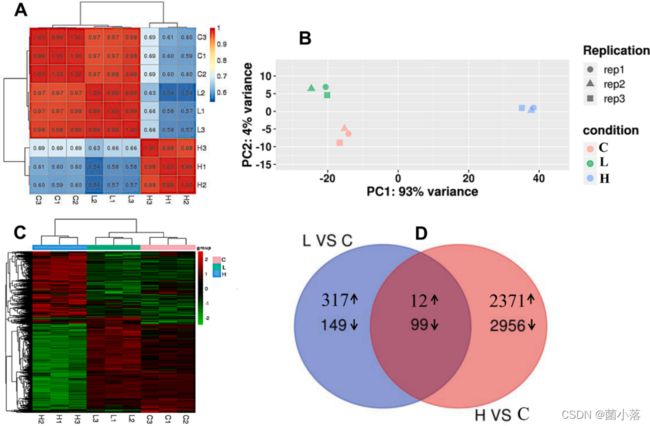
图2 不同浓度的TYN处理后,羊角月牙藻的转录组分析
3.GO、KEGG富集分析
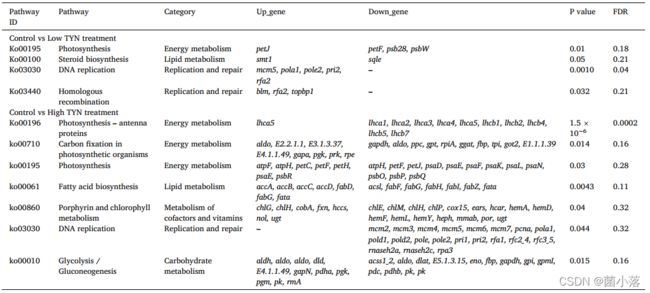
在GO分析中,低、高TYN处理组分别富集了720和3095个GO条目。KEGG代谢途径分析显示,在低TYN处理组,DNA复制、同源重组和光合作用等8条途径受到显著影响;相比之下,高TYN处理组发生改变的15条通路中,主要为光合天线蛋白、DNA复制、脂肪酸生物合成、光合生物体固碳和光合作用等(表1)。
表1 TYN处理后,各处理组与对照组相比显著富集的代谢通路
4.代谢组学分析
在正离子和负离子模式下分别检测出355和415个代谢物。与对照组相比,在负离子模式下,低、高处理组分别鉴定出57和84个差异代谢物;在正离子模式下,低、高处理组分别鉴定出40和94个差异代谢物。
对所有差异代谢物合并进行KEGG富集分析表明,低、高处理组分别富集到5个和6个显著差异代谢通路。在两个处理组中代谢途径,次生代谢物的生物合成,细菌分泌系统和光合作用都显著富集。
5.转录组学和代谢组学联合分析
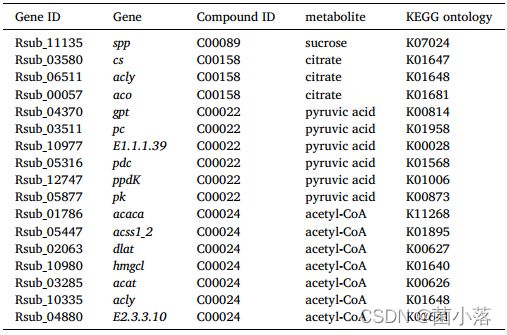
对转录本和代谢产物的综合分析表明,与对照组相比,低、高处理组共发现18个和52个差异代谢物与至少一个转录本配对(表2)。例如,在低TYN处理组中,差异代谢物及其相关基因腺苷(cmpk1)、磷酸(ap1_2)、腺苷酸(E3.1.3.5)、棉子糖(E2.4.1.82)等在嘧啶代谢、嘌呤代谢、半乳糖代谢等通路中均有相同的上下表达趋势。而在高TYN处理组中,磷酸(metK)、丙酮酸(ppdK)、乙酰辅酶a (dld)、尿嘧啶(psuG)等在半胱氨酸和蛋氨酸代谢、光合生物体固碳作用、柠檬酸循环、嘧啶代谢等代谢通路中表达趋势相同。
表2 处理组中差异代谢物与相应转录本的关联
讨论
本研究揭示了TYN对羊角月牙藻的毒性分子机制。与我们的假设一致的是,在低和高TYN处理组中,TYN显著改变了与DNA复制、光合作用和能量生产相关的分子通路。在TYN及其衍生物的长期胁迫下,暴露于TYN下的藻类生长抑制可能是由于光合作用抑制、诱导氧化应激、DNA复制耦合修复增加和能量代谢失调等原因导致的。
1.藻类在TYN胁迫下解毒的相关基因
在藻类中,外源性化合物代谢包括生物转化(阶段I和阶段II)和运输(阶段III)。在本研究中,TYN处理后,参与这三个解毒阶段的基因在羊角月牙藻中大多下调。在I期,细胞色素P450 (Cyt P450)在低TYN和高TYN处理组均下调;此外,乙醇脱氢酶(adhs)、醛脱氢酶(aldhs)和短链脱氢酶(sdrs)仅在高TYN处理组下调。这些结果表明,高TYN处理组的解毒过程可能受到抑制。本研究中,低TYN处理组ABC转运蛋白家族A、G基因表达显著上调,而在高TYN处理组中显著下调。因此,在低TYN处理组中,这些基因的上调可能有助于TYN从藻类细胞中消除,降低TYN暴露引起的毒性;然而,在暴露于高水平TYN处理组中,这些基因的下调显著抑制了羊角月牙藻的生长。
2.与光合作用相关的分子途径
本研究中,高TYN处理组中光合作用天线蛋白、卟啉和叶绿素代谢以及光合生物的碳固定等信号通路受到显著影响。在低TYN和高TYN处理组中,光合作用信号通路均发生了显著变化。这些结果表明,高水平的TYN处理可能抑制了藻类叶绿素的生物合成和光采集信号。由于TYN诱导的光合抑制,活性氧(ROS)的过度产生可能导致氧化应激和生长抑制。参与C4循环过程的基因(ppc, got2, gpt, E1.1.1.39)下调,表明本研究中,在高TYN胁迫下,CO2的释放可能减少。在低和高TYN处理组中,丙酮酸、磷酸和ATP含量在代谢水平下降,海藻生长受到抑制。
3.与DNA复制耦合修复相关的分子途径
在本研究中,参与DNA复制过程的20个DEGs在高TYN处理组中持续下调,这一生物学过程可能是TYN导致藻类生长性能下降的原因(表1)。这些被下调的DEGs编码的蛋白质参与了DNA复制的不同阶段。编码核糖核酸酶H2的rnaseh2a和rnaseh2c的下调表明DNA复制的终止信号被抑制(表1)。DNA复制耦合修复的抑制可能导致细胞分裂的减少,DNA损伤的积累,以及潜在的基因组不稳定,这表明羊角月牙藻在高TYN处理组的生长性能下降。此外,代谢组学水平上ATP含量的降低可能与TYN胁迫下羊角月牙藻的DNA复制和修复有关。
4.与能量代谢相关的分子途径
本研究中,高TYN处理组的半乳糖代谢、糖酵解等碳水化合物代谢相关通路受到显著影响(表1)。在代谢水平上,两组TYN处理组的柠檬酸、正磷酸和ATP含量均明显下降。相比之下,高TYN处理组中乌头酸、丙酮酸和蔗糖含量降低,乙酰辅酶a含量增加。蔗糖、丙酮酸、柠檬酸和ATP含量的降低,暗示了烃类代谢的失调和能量生产的减少,从而导致藻类生长受到抑制。乙酰辅酶a作为烃类代谢和脂类代谢之间的关键中间体,进入柠檬酸循环并被氧化生成能量。因此,丙二酰辅酶a的积累表明,在高TYN处理组中,TCA循环的阻塞或脂肪酸氧化的增加。脂质代谢的失调可能导致ROS的过度产生、氧化应激的诱导和藻类生长的抑制。
文章小结
3μg L-1和400μg L-1的TYN影响了羊角月牙藻光合作用、DNA复制耦合修复和能量代谢相关的分子途径。与TYN在细菌中的作用模式(TYN与50S核糖体结合,抑制蛋白质合成)相反,光合作用信号通路是TYN对于羊角月牙藻毒性最敏感的靶点之一。而在高TYN处理中,生长抑制与光合系统、DNA复制耦合修复、烃类代谢、脂质代谢和氧化应激相关的分子途径有关。值得注意的是,即使在环境中浓度较低的情况下,TYN的持续存在也可能对初级生产者(如藻类)和食物链中较高营养水平的消费者造成不良影响。