易基因|组学研究:人类胎儿发育过程的DNA羟甲基化(5hmC)和RNA m5C动态重编程
易基因|组学研究:人类胎儿发育过程的DNA羟甲基化(5hmC)和RNA m5C动态重编程
大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
2022年5月26日,中国科学院研究团队合作以“Dynamic DNA 5-hydroxylmethylcytosine and RNA 5-methycytosine reprogramming during early human development”为题在权威学术期刊《Genomics Proteomics & Bioinformatics》(IF 7.691)发表研究论文,该研究通过微量DNA羟甲基化测序技术5hmC-Seal、RNA m5C甲基化检测技术RNA-BS和对应的转录组RNA-seq等组学测序结果揭示了人类胎儿发育过程的DNA 5-羟甲基胞嘧啶(5hmC)和 RNA 5-甲基胞嘧啶(m5C)动态重编程机制。

标题:Dynamic DNA 5-hydroxylmethylcytosine and RNA 5-methycytosine reprogramming during early human development
发表时间:2022年5月26日
期刊:Genomics Proteomics & Bioinformatics
影响因子:IF 7.691
技术方法:微量细胞DNA羟甲基化测序(5hmC-Seal)、 RNA重亚硫酸盐测序(RNA-BS)、转录组测序(RNA-seq)等(易基因优势技术)
研究摘要:
人胚胎植入后,复杂和高度专业化的分子事件呈现功能上不同的器官形成,而表观基因组如何调控器官特异性发育仍有待阐明。本研究纳入123个人13-28周胎儿的的心脏、肾脏、肝脏、肺器官样本,通过微量5hmC-Seal、RNA-BS和RNA-seq等测序技术绘制了DNA 5hmC和RNA m5C表观基因组图谱。分别鉴定出70091个器官阶段特异性差异羟甲基化区域(DhMR)和503个mRNA m5C修饰。发现关键转录因子(TFs)、T-box转录因子20(TBX20)、配对盒蛋白pax-8(PAX-8)、krueppel样因子1(KLF1)、转录因子21(TCF21)和CCAAT增强子结合蛋白β(CEBPB)特别有助于在不同阶段形成不同的器官。此外,分析结果还发现5hmC富集的Alu元件可以参与TF靶向基因表达的调节。本研究揭示了DNA修饰和RNA甲基化之间的重要联系,并说明了人类胎儿器官发生过程中的表观遗传图谱,为理解早期发育和出生缺陷的深入表观遗传机制提供了基础。
研究背景:
表观遗传修饰包括DNA甲基化、组蛋白修饰、染色质可及性、RNA修饰等。它不仅作为染色质组织和结构规范的可遗传和稳定指示,而且还参与哺乳动物胚胎发育过程中转录状态的调节。人类胚胎和成人器官在生理和病理条件下的表观基因组已被广泛研究;然而,与人类胎儿器官正常发育有关的表观遗传网络仍不清楚。
方法:
在胎龄13-28周选择性引产后的胎儿中获得123个胎儿心脏、肝脏、肾脏和肺器官组织样本(排除病理妊娠、异常核型或发育缺陷的胎儿组织)。解剖后分离所有正常器官,用医用盐水连续冲洗3次,立即液氮冷冻。随后进行基因组DNA和RNA制备,用于后续的微量5hmC-Seal、RNA-BS和RNA-seq测序分析。
结果:
(1)人类胎儿器官全基因组DNA 5hmC分析
作者分别在13-15、18-19、21-23 和 25-28 周的 8 个人类胎儿中收集了心脏、肾脏、肝脏、肺 4 个主要器官的样本,每个阶段重复 2-4 次。为研究全基因组 DNA 5hmC 特征,进行了微量细胞DNA羟甲基化测序(5hmC-Seal)。

图1:早期人类胎儿发育的DNA 5hmC图谱
A.人类胎儿器官的发育时间表和本研究中使用的器官样本和策略示意图。
B.器官特异性基因(TRDN,PAX2,RASA4B和FAM86FP)的IGV视图。
C.DNA 5hmC在心脏,肾脏,肝脏和肺组织不同发育阶段的Metagene图。
D.相对于不同器官中预期的DhMRs和ChMRs在不同基因组区域的归一化富集得分,正值表明富集程度超过预期。
E.DhMRs在不同器官转录起始位点(TSS)的分布。
F.不同器官样品的器官和阶段特异性DhMR中5hmC信号主成分分析(PCA)。
G.器官和阶段特异性DhMR的5hmC信号热图。
H.器官和阶段特异性DhMR的GREAT分析结果。
(2)人类胎儿器官发育过程中的DNA 5hmC重编程
5hmC 动态变化分析结果表明,转录因子(TF)可能在胎儿5hmC水平动态序列变化的帮助下参与特定器官的发育。表明DNA 5hmC可以作为表观遗传标记 Alu 元件的过渡状态,通过5hmC影响 TF 结合和下游基因表达。

图2:DNA 5hmC通过TFs和TEs动态促进胎儿器官发育
A.心脏、肾脏、肝脏和肺部器官特异性DhMRs的K均值聚类分析,每个器官分为6个簇。
B.DhMRs鉴定的TF motifs热图。
C.胎儿肾脏器官和阶段特异性DhMRs中特定TE家族的富集分析。
D.簇3和簇4中Alu元件CEBPB结合位点的富集分数,与所有Alu元件CEBPB结合位点的总富集(红色)及其全基因组分布(蓝色)相比较条形图。条形图还显示所有Alu元件所有CEBPB结合位点与其全基因组分布相比的富集情况(灰色)。
E.将D中CEBPB靶向Alu元件的5hmC信号和下游基因表达水平z评分标准化。
(3)DNA 5hmC与人体器官的转录组稳态相关
大多数DhMRs位于所有胎儿器官的TSS区域周围(图 1E)和DhMRs上5hmC信号的序列变化表明,在器官发育过程中,5hmC信号与RNA丰度密切相关(图 2B和E)。因此,研究人员分析启动子上的DNA 5hmC动态重编程是否在发育阶段调控基因表达。分析结果发现启动子区域的5hmC修饰与胎儿器官发育期间的基因表达稳态高度相关。
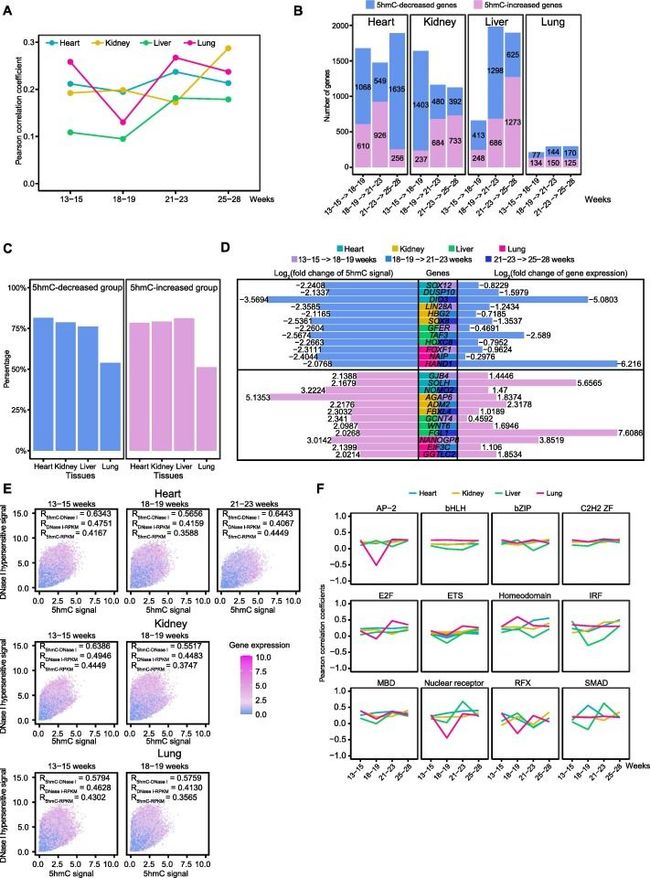
图3:DNA 5hmC参与调控人类胎儿器官的基因表达
A.启动子不同阶段5hmC信号与基因表达水平的Pearson相关系数中位数。
B.胎儿器官发育过程中5hmC显著减少和增加的基因数量。
C.每个器官中5hmC显著变化的器官特异性基因的百分比。
D.5hmC信号的倍数变化(log2)和5hmC减少组/增加组18个器官特异性基因的基因表达水平。
E.双标图(Biplots)比较5hmC信号(x轴)、DNase I超敏信号(y轴)的变化与RNA丰度变化(颜色)。每个点代表一个基因。
F.心、肾、肝、肺不同TF家族组pearson相关系数的动态变化。基于5hmC信号和相应基因组中的基因表达水平计算Pearson相关系数。
(4)RNA m5C在胎儿器官发生过程的下游起调控作用
RNA m5C是与RNA稳定性相关的普遍RNA转录后修饰之一,研究人员对13-28周三个时期的心、肾、肝、肺四个胎儿器官进行RNA重亚硫酸氢盐测序(RNA-BS)。结果揭示了在全基因组范围内,m5C修饰水平在肾、肝和肺的发育过程中动态变化。进一步结果表明,RNA m5C在人类胎儿器官发育过程中充当转录后调节器。

图4:胎儿器官发育过程中的RNA m5C甲基化图谱
A.心脏、肾脏、肝脏和肺不同样本中不同区域的m5C分布。
B.m5C位点和m5C修饰mRNAs数量。
C.心脏、肾脏、肝脏和肺不同发育阶段的总RNA m5C甲基化水平。
D.通过比较相邻阶段样本的RNA序列数据,显示m5C修饰和m5C不修饰的mRNAs表达水平变化累积分布。
E.器官和阶段特异性基因的m5C甲基化水平与胎儿器官发育过程中相应基因的表达水平之间的强正相关热图。
(5)DNA 5hmC和RNA m5C的特异性和协同作用
接下来,研究人员提出一种可能:DNA 5hmC和RNA m5C可能相互作用,在时空上调节胎儿器官的调控网络。为进一步研究DNA 5hmC和RNA m5C的各自功能,研究人员将器官特异性修饰基因分为4组:由DNA 5hmC和RNA m5C共同特异性调节的差异表达基因(A组)、仅由DNA 5hmC特异性调节的差异表达基因(B组)、仅由RNA m5C特异性调节的差异表达基因(C组)、由DNA 5hmC和RNA m5C特异性调节的常见表达基因(D组)。
分析结果发现,在A组中,DNA 5hmC信号强度、RNA m5C甲基化水平和基因表达水平之间存在密切关联。在胎儿肾脏和肝脏中,DNA 5hmC和RNA m5C可能表现出不同的特异性功能。进一步结果表明,DNA 5hmC和RNA m5C可能分别通过转录前和转录后机制促进器官特异性调控网络,这些机制由TET3和NSUN2/ALYREF介导。关于如何在机制上促进发展过程,仍值得进一步详细研究。

图5:DNA 5hmC和RNA m5C持续调节胎儿器官发育
A.DNA 5hmC-RNA m5C共调节特异表达基因(A组)的启动子DNA 5hmC信号(左)、m5C甲基化水平(中)和表达水平(右)动态变化热图。DNA 5hmC信号、m5C甲基化水平和基因表达水平通过z评分标准化。
B.A组、B组和C组中器官特异性基因的GO生物过程富集分析。
C.m5C修饰或不修饰相应基因启动子上的5hmC信号。
D.胎儿器官中5hmC甲基转移酶TET3和m5C“writer”NSUN2、“reader”ALYREF和YBX1的表达水平之间相关性。
结论:
综上所述,这项研究不仅揭示了植入后胚胎器官发生过程中DNA 5hmC和RNA m5C的动态重编程,而且还揭示了DNA修饰和RNA甲基化之间的相互作用。通过转录组可以时空协同调节,在特定发育阶段形成协同良好的网络,确保胎儿正常发育。此外,对DNA和RNA修饰水平的特征和功能重要性的全面描述提供了可用的人类胎儿表观基因组数据集,并为深入理解人类早期发育的表观遗传学机制奠定了基础。

关于DNA羟甲基化
羟甲基化5hmC是哺乳动物基因组上的第六碱基,在发育、衰老、神经退行性疾病、复杂疾病及肿瘤发生过程中起重要作用。DNA羟甲基化是近年发现的一种新的DNA修饰并迅速成为研究热点。随着研究的深入,发现之前被认为是检测DNA甲基化标准的重亚硫酸盐测序并不能区分DNA甲基化(5mC)和DNA羟甲基化(5hmC)。
传统BS转化无法区分5mC和5hmC
传统的Bisulfite测序中,5hmC经过Bisulfite处理后变为CMS,CMS在测序中仍然被读作C碱基,因此不能区分5mC和5hmC。

易基因联合剑桥大学建立了化学氧化法结合重亚硫酸盐转化的测序技术(oxidative bisulfite sequencing, oxBS-Seq),该技术不仅可以精确检测DNA甲基化,排除DNA羟甲基化的影响,还可以双文库结合同时单碱基分辨率精确检测DNA羟甲基化。
易基因羟甲基化测序技术满足多种应用需求:
- 全基因组氧化甲基化测序(oxWGBS)
- 简化基因组氧化甲基化测序(oxRRBS)
- 目标区域靶基因氧化甲基化测序(Target-oxBS)
- 羟甲基化DNA免疫沉淀测序(hMeDIP-seq/5hmC-Seal)
关于RNA m5C甲基化
m5C是RNA百余种修饰中研究较多的一种。m5C存在于tRNA上时,可以对翻译进行调节;存在于rRNA上时,可以对核糖体的生物合成进行质控;存在于mRNA上时,则可以影响mRNA的结构、稳定性及翻译过程。
易基因提供适用于不同科研需求的m5C甲基化测序技术:
- 常规mRNA m5C甲基化测序(RNA-BS):
mRNA分离后首先通过亚硫酸盐处理,非甲基化的C转变为U,m5C修饰的碱基保持不变,结合高通量测序,可以对RNA上的每一个C碱基修饰进行定位与定量。
- 常规mRNA +lncRNA m5C甲基化测序(全转录组RNA-BS):
易基因科技建立的升级版m5C RNA甲基化测序服务,去除人rRNA后,剩余RNA经重亚硫酸盐处理后,结合高通量NGS策略,可在全转录组范围内单碱基分辨率地检测基因m5C甲基化修饰分布。

技术优势:
- 高深度:超高深度重亚硫酸盐处理,检测准确性极高;
- 高通量:结合高通量NGS,全转录组范围内检测;
- 单碱基:单碱基分辨率,快速检测和分析RNA中的m5C。
- 高准确:精确的检测mRNA等每一个C碱基的的修饰水平。
易基因技术团队多组学关联分析方法总结
1. 直接关联
一个基因的功能元件甲基化情况影响该基因的表达。
• 重叠分析
• Pearson/Spearman 相关性分析
2. 模型关联
基于基因转录、蛋白质、代谢物等之间的上下游相互作用联系。
• 多元线性模型(multiple linear model)
3. 网络关联
基于分子功能和通路的富集性。
• WGCNA module correlation
• EMDN algorithm
• SNF algorithm
从关联走向因果:组学分子实验验证
基因表达相关的组学:
- 基因敲除/抑制
- 基因过表达
甲基化组学:
- 甲基化酶基因的敲除与过表达
宏基因组(肠道菌群):
- 无菌动物模型
- 粪菌移植
参考文献:
Han X,et al. Dynamic DNA 5-hydroxylmethylcytosine and RNA 5-methycytosine reprogramming during early human development. Genomics Proteomics Bioinformatics. 2022 May 26.
更多阅读:
项目集锦 | 易基因DNA羟甲基化5hmC测序研究成果
项目文章|DNA(羟)甲基化研究揭示铁离子依赖表观调控促进狼疮致病性T细胞分化
综述:如何研究植物基因组DNA羟甲基化(5hmC)?
表观技术 | 靶基因DNA甲基化和羟甲基化测序 定制精准检测
专注表观组学十余年-深圳市易基因科技有限公司