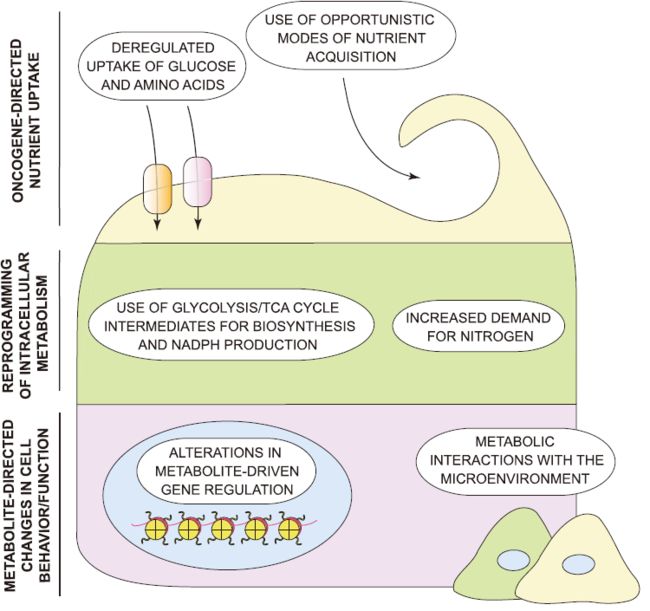
癌症代谢标志[1]
与肿瘤发生有关的细胞代谢可分为三个阶段六个标志(图1):
- 通过赋予获得所需营养的能力增强来影响代谢物流入
- 葡萄糖和氨基酸的摄取失调
- 营养获取的适应性模式
- 塑造营养物质优先分配给有助于细胞致瘤特性的代谢途径的方式
- 糖酵解/TCA循环中间产物用于生物合成和NADPH产生
- 对氮的需求增加
- 对细胞命运产生长远影响,其中包括癌细胞本身以及肿瘤微环境组成的分化改变
- 代谢物驱动的基因调控改变
- 代谢与肿瘤微环境相互作用
1. 癌细胞中葡萄糖和氨基酸的摄取失调
细胞中葡萄糖和谷氨酰胺的作用:
为了满足与增殖有关的生物合成需求,细胞必须增加从环境中吸收养分。支持哺乳动物细胞存活和生物合成的两种主要营养物质是葡萄糖和谷氨酰胺。
- 通过葡萄糖和谷氨酰胺的分解代谢,细胞可以维持多种碳中间产物的浓度,这些碳中间产物被用作组装各种大分子的基础。
- 葡萄糖和谷氨酰胺碳骨架的受控氧化使细胞能够以NADH和FADH2的形式捕获其还原能力,后者介导电子向电子传输链的转移以促进ATP的产生,或以相关的辅助因子NADPH,它可以降低多种生物合成反应的能量,并有助于维持细胞的氧化还原能力。
- 谷氨酰胺是一种含有两个还原氮原子的非必需氨基酸,是还原氮在细胞间运输的主要方式,同时谷氨酰胺的酰胺基是嘌呤和嘧啶碱生物合成所必需的硝基供体,因此谷氨酰胺提供了嘌呤和嘧啶核苷酸,6-磷酸氨基葡萄糖和非必需氨基酸的生物合成所需的氮。
- 用于尿嘧啶和胸腺嘧啶的生产需要一分子谷氨酰胺,胞嘧啶和腺嘌呤各需要两分子,而建立一个鸟嘌呤碱需要三分子谷氨酰胺。
- 嘧啶环和嘌呤环的组装都利用天冬氨酸,天冬氨酸来自TCA循环代谢物草酰乙酸和谷氨酸的转氨作用,两者都是谷氨酰胺的分解代谢产物。
- 利用谷氨酰胺的酰胺氮,天冬酰胺合成酶(ASNs)催化天冬氨酸合成天冬酰胺。天冬酰胺是哺乳动物细胞唯一不分解的氨基酸。同时天冬酰胺在谷氨酰胺缺乏的情况下发挥着关键的调节作用。
- 谷氨酰胺可以被谷氨酰胺酶直接脱酰胺为谷氨酸。谷氨酸是氮的供体,通过转氨作用产生许多非必需氨基酸。
- 谷氨酰胺在摄取必需氨基酸中也起作用。通过质膜定位的中性氨基酸反转运蛋白LAT1导入必需氨基酸亮氨酸与释放谷氨酰胺有关。因此,细胞内谷氨酰胺可以促进LAT1底物的摄入,包括亮氨酸,异亮氨酸,缬氨酸,蛋氨酸,酪氨酸,色氨酸和苯丙氨酸。
90多年前德国生理学家Otto Warburg首次描述与不扩散的正常组织相比,肿瘤对葡萄糖的消耗明显增加。该观察结果已在多种肿瘤背景下得到证实,并与不良的肿瘤预后相关。
1950年代,美国生理学家哈里·埃格首先描述了肿瘤细胞对谷氨酰胺增殖的高需求,他证明培养的HeLa细胞的最佳生长需要相对于其他培养基在培养基中的谷氨酰胺过量10到100倍摩尔氨基酸。
此外,发现谷氨酰胺是艾氏腹水癌以及体内大量增殖的肝癌和肉瘤肉瘤中消耗最快的氨基酸。
与相应的正常组织相比,许多致瘤背景与肿瘤环境中谷氨酰胺的消耗有关。
什么导致肿瘤细胞内化大量葡萄糖和谷氨酰胺?
正常细胞的营养摄入受严格调控:
- 后生动物尽管被富含营养的血浆和细胞外液所包围,但不会以组成型方式摄入营养。营养的吸收受到生长因子信号的严格调控。尽管培养基中葡萄糖的含量很高,当缺乏生长因子时,造血和神经细胞无法消耗足够数量的葡萄糖,甚至无法维持细胞生物能。然而,通过与质膜葡萄糖转运蛋白GLUT1和糖酵解途径己糖激酶(HK)结合表达,可以很容易地恢复缺乏生长因子的细胞的存活。
- 除可溶性生长因子外,细胞与细胞外基质的相互作用在调节葡萄糖摄取中也起作用。在细胞与细胞外基质分离的条件下培养乳腺上皮细胞会损害葡萄糖的吸收并导致线粒体电位降低和ATP水平降低。综上所述,这些观察结果表明,葡萄糖的摄入不是由细胞的即时生物能需求驱动的,相反,葡萄糖的分配是严格定量的。
为了避免缺乏代谢资源,异常激活的癌基因和/或肿瘤抑制因子的丧失使癌细胞维持在一种从细胞外环境组成性摄取可用葡萄糖,谷氨酰胺和必需氨基酸的状态。 与需要粘附和生长因子驱动的信号输入来维持生存和增殖的正常细胞相反,癌细胞积累的致癌性改变使其独立于这些外部需求。特别是靶向PI-3激酶的负调节剂PTEN和INPP4B的遗传改变以及多种上游受体酪氨酸激酶中的激活突变和基因扩增会导致多种癌症类型的组成型葡萄糖摄取和代谢。
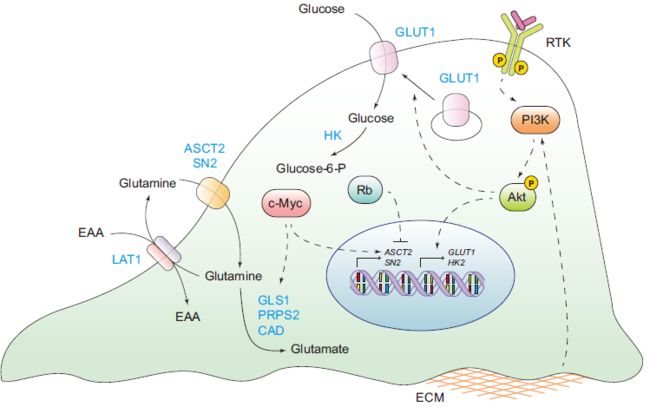
癌症中促进细胞获取营养的生长信号(图2)
-
PI3K/Akt信号传导是受体酪氨酸激酶以及细胞外基质的信号的汇聚点,是葡萄糖摄取的主要调节剂。
- PI3K/Akt信号传导既促进葡萄糖转运蛋白GLUT1 mRNA的表达,又促进GLUT1蛋白从内膜到细胞表面的转运。
- Akt增强了HK(使葡萄糖分子磷酸化,阻止了它们回流到细胞外空间)以及磷酸果糖激酶(PFK,催化了糖酵解的关键不可逆步骤)的活性。
- 肿瘤中的18F-FDG-PET信号强度与PI3K/Akt通路活性水平密切相关,并被PI-3激酶和受体酪氨酸激酶抑制剂减弱。
- 活性形式的Akt的外源表达可刺激糖酵解,并足以恢复生长因子缺乏细胞中的细胞大小,活力,线粒体电位和ATP水平。
- Akt的组成型活性形式还可以防止细胞附着丧失引起的ATP水平下降。
- 在生物合成需求增加的生理环境中,Akt信号传导对促进葡萄糖摄取至关重要。例如,小鼠乳腺中Akt1的靶向缺失消除了泌乳诱导的葡萄糖摄取增加,从而导致乳汁生产不足。
Ras,可以上调GLUT1 mRNA的表达并增加细胞葡萄糖的消耗。
-
转录因子c-myc在增殖细胞中上调,并经常在各种肿瘤类型中被扩增靶向,是增殖细胞利用谷氨酰胺的主要驱动力。
- c-myc诱导谷氨酰胺转运蛋白ASCT2和SN2的转录,促进谷氨酰胺酶(GLS1),磷酸核糖焦磷酸合成酶(PRPS2)和氨甲酰磷酸合成酶2(CAD)的表达。
- 通过将谷氨酰胺转化为谷氨酸来支持转运蛋白促进的谷氨酰胺摄取。产生的谷氨酸不能通过谷氨酰胺转运而离开细胞,并且随着其积累,通过充当半胱氨酸反转运蛋白xCT的交换底物,促进TCA周期异常并刺激半胱氨酸的摄取。
除了c-myc的正调控外,Rb肿瘤抑制蛋白家族还对谷氨酰胺的摄取进行负调控。Rb家族蛋白的缺失通过E2F依赖性的AS2和GLS1上调来上调谷氨酰胺的摄取和利用。细胞分裂的两个关键促进因子c-myc和E2F发挥作用的部分原因是使谷氨酰胺能够进入细胞。
2. 癌细胞营养获取的适应性模式
尽管具有吸收葡萄糖和氨基酸的亲和力,但由于营养物消耗速率的增加和肿瘤血管供应的不足,体内癌细胞经常会遇到营养物缺乏的状况。肿瘤细胞利用适应性的营养获取模式,从而使其能够在代谢不利的条件下生存和增殖。这些适应包括:
1. 获取通常无法获得的营养来源。
2. 当预制分子在细胞内的合成受到损害时回收预制分子的能力。
3. 即使没有可用的细胞外营养,也可利用适应性代谢途径维持其生存。
获取通常无法获得的营养来源
-
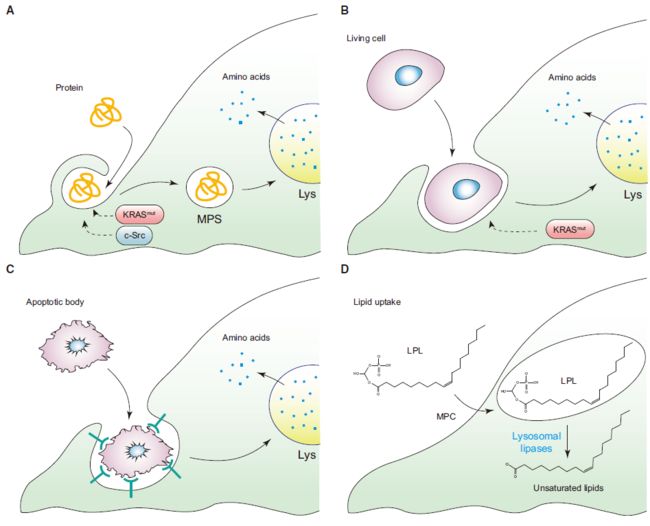
组织的血浆和间质液富含可溶性蛋白,但通常不被用作氨基酸来源。突变Ras或c-Src等位基因的表达使细胞能够通过胞外蛋白的溶酶体降解来恢复游离氨基酸(图3A)。
- 与通过专门的转运蛋白进入细胞的低分子量营养摄取相反,胞外大分子的捕获通过巨胞饮作用将大量胞外液吸收到巨大的囊泡中。Ras和c-Src驱动的肌动蛋白细胞骨架重塑刺激巨胞饮作用。充满液体的巨胞体被运输到细胞内部与溶酶体融合,被吞噬的蛋白质被蛋白酶水解,释放出游离氨基酸。
- 在生理水平(20-30 mg / mL)的血清白蛋白上添加培养基可以使KRASG12D转化的MEF在缺少必需氨基酸亮氨酸的情况下增殖。已经证明补充白蛋白可以在不含所有游离氨基酸的培养基中恢复KRASG12D驱动的小鼠胰腺癌系的增殖。
- mTORC1抑制剂可大大提高细胞从捕获的细胞外蛋白中回收氨基酸的能力,并在缺乏必需氨基酸的情况下增强其生长。在氨基酸充足的条件下,mTORC1抑制了细胞外蛋白质作为营养物的利用,仅在游离氨基酸不足的情况下用作应急来源。
除可溶性细胞外蛋白外,癌细胞还可以通过吞噬和吞噬整个活细胞来回收游离氨基酸(图3B),以及从凋亡细胞的吞噬作用中回收游离氨基酸(图3C)。在此过程中带有突变KRAS等位基因的细胞更容易发生内亡。这种性质可能有助于肿瘤内的细胞竞争,从而导致更具侵略性的细胞群体的出现。
内亡(Entosis)或叫侵入性细胞死亡,是一个新的细胞死亡方式,其中一个细胞吞噬另一个活细胞,被吞噬细胞最后死于吞噬小体。在侵入性死亡过程中,细胞通过钙黏着蛋白介导的细胞间连接作用,经内化过程侵入临近的宿主细胞空泡内,被溶胞体酶系降解而导致死亡。
利用适应性代谢途径维持其生存
- 癌细胞可以通过巨自噬的自分解过程承受长期的营养剥夺。在自噬过程中,细胞内大分子和整个细胞器被双膜结构包裹,并与溶酶体融合。一旦进入溶酶体,被吞噬的货物就会被常驻的蛋白酶和脂肪酶降解,从而释放出游离的氨基酸和脂肪酸。自噬可以使营养或生长因子缺乏的细胞在培养中维持其活力数周。但是,自噬不能为细胞提供新的生物质,因此不能在营养缺乏的情况下支持增殖。
当预制分子在细胞内的合成受到损害时回收预制分子的能力
在生长中的肿瘤中缺乏血管营养输送的第二个结果是缺氧区域的出现导致抑制了许多需要分子氧作为电子受体的生物合成反应。缺氧损害了硬脂酰辅酶A去饱和酶1(SCD1)催化的从头产生的脂肪酸中双键的引入,造成了不饱和脂肪酸种类的不足。
- 为了补充缺少的脂肪酸,低氧细胞以单个含酰基链的溶血磷脂的形式从周围环境中导入“现成的”不饱和脂肪酸(图3D)。因此,当在低氧条件下培养时,去除血清脂质会在某些癌细胞中触发内质网应激和凋亡。
- 在其他癌细胞中,更复杂的游离脂肪酸的胞外释放增加。在多种癌症亚型中检测到脂蛋白脂肪酶和单酰基甘油脂肪酶(MAGL)的表达升高,并且与侵袭性相关。
- 某些肿瘤的适应能力强,可以使他们直接从血浆中回收脂肪酸,但其他肿瘤则可以诱导邻近的正常细胞释放脂质。例如,转移性卵巢癌细胞表面上长链脂肪酸结合蛋白FABP4的上调使它们可以直接从网膜脂肪的脂肪细胞中获取脂肪酸。
3. 糖酵解、TCA循环中间产物用于生物合成
- 细胞增殖不仅增加细胞需要的营养物还改变营养的使用方式。
- 当肿瘤细胞是不增殖时,葡萄糖被优先地用于线粒体的乙酰辅酶A产生,然后将其进行氧化的TCA循环。从TCA循环的氧化反应中提取的电子通过NAD+/NADH和FAD/FADH2传递给电子传递链,产生的电化学梯度介导ATP产生。
- 增殖细胞的碳代谢与非增殖的细胞显着不同。在增殖细胞中碳的主要是用于生物分子的多样性阵列的生物合成,其中大量的生物合成是脂肪酸和胆固醇,戊糖和己糖衍生物,甘油,核苷酸和非必需氨基酸。
- 为了实现细胞增殖,增殖细胞必须首先转换获取的营养物质为不同的中间产物(图4B),包括胞质乙酰-CoA,一碳载叶酸循环单元和S-腺苷甲硫氨酸(SAM),以及糖酵解和TCA循环中间产物。
- 许多生物合成反应具有还原性,因此需要还原力。例如,代棕榈酸的消耗需要14还原当量,同时建立一个胆固醇分子需要26。在细胞的生物合成反应中使用的还原当量的指定供体是NADPH。从NADP+产生NADPH是由碳底物受控氧化产生的,但不同于产生NADH的途径。因此,增殖细胞必须分配它的一部分碳底物,用于NADPH的生产。
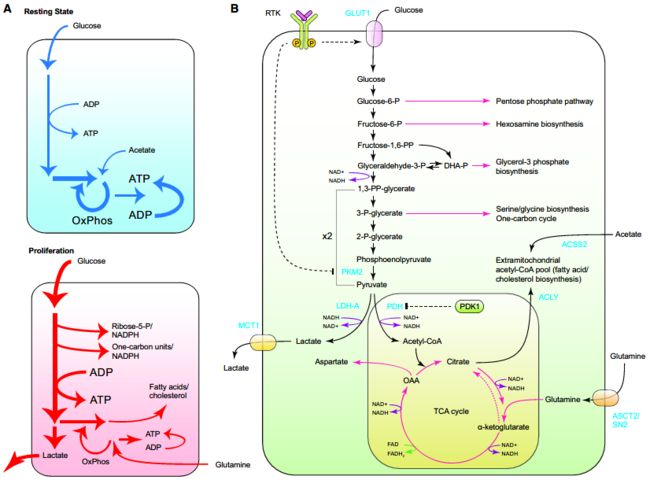
Warburg效应
碳代谢由增殖细胞重编程,尽管消耗葡萄糖大量的并且在暴露于环境氧气,细胞没有采取糖酵解与TCA循环耦合所提供的生物能量优势。相反,在富含葡萄糖的培养基中生长时,细胞转换剩余丙酮酸为乳酸,并分泌到细胞外环境中,这让肿瘤细胞通过一个相当低效的途径从糖酵解的底物水平磷酸化反应中获得ATP。
然而,肿瘤细胞具有功能性线粒体和保留以进行氧化磷酸化的能力。事实上,靶向性缺失线粒体DNA减少了癌细胞系的致瘤潜力在体外和体内。此外,优先转化葡萄糖为乳酸已被证实存在于遗传正常增殖细胞以及在由病毒感染的细胞。
- 细胞增殖所需的线粒体电子传输链(ETC)的关键功能是能够合成天冬氨酸(一种蛋白质原氨基酸,嘌呤的前体)和嘧啶合成。天门冬氨酸因ETC活性缺陷受到抑制。
- 丙酮酸通过乳酸脱氢酶在细胞质中再生NAD+ 。该NAD+可以激活细胞质苹果酸脱氢酶,从苹果酸中提供OAA,并通过GOT1(编码胞质天冬氨酸转氨酶)驱动天冬氨酸的合成逆转了ETC活性缺陷的抗增殖作用。[2]
与肿瘤的增殖细胞相比,肿瘤细胞的静态亚群明显减少糖酵解,并表现出更高的依赖性氧化磷酸化,以及线粒体呼吸组分的高表达。这些结果表明,Warburg效应是一个被调节的代谢状态,并且在生物合成需求增加的时期是有益的。
多种癌基因已被证明有助于增殖细胞的代谢适应。c-myc协同增加PDK11的表达。b-catenin/tcf信号也被证明上调MCT1和PDK1转录。HIF1A通过缺氧或在不同致癌背景下的稳定也触发LDH-A和PDK1的协调转录上调。
乳酸脱氢酶a(LDH-A)是一种催化丙酮酸还原为乳酸的酶;单羧酸转运体(MCT1)是一种促进乳酸进入细胞外空间的转运体。
为什么增殖细胞要将过量的丙酮酸转换为乳酸,而不是保持氧化磷酸化?
相对于ATP相关的前体分子和NADPH形式的还原当量,增殖细胞只略微增加它们所需要消耗的ATP。葡萄糖代谢是这些前体和还原当量的稳定提供者,而活化的TCA循环产生的NADH和ATP是葡萄糖代谢的主要负调节物。
通过将过量的丙酮酸转化为乳酸,增殖细胞预防胞浆中NADH的积聚并减少ATP产生,通过促进连续的胞质葡萄糖代谢解除过量的线粒体ATP产生带来的负反馈。
-
过量的糖酵解流被优先变换为乳酸,以帮助保存足够的NAD+维持糖酵解并避免线粒体中NADH过量供给造成TCA循环抑制。
- 糖酵解产生的过量NADH并不是通过化学穿梭导入增殖细胞的线粒体,而是利用定位于线粒体外膜上的甘油磷酸脱氢酶(MGPDH)将糖酵解中间产物DHAP转化为甘油-3-磷酸,甘油-3-磷酸直接向电子传递链提供电子。
- 通过使用FAD作为电子受体,mGPDH催化反应降低了通过糖酵解产生NADH带来的线粒体净电化学势。
-
线粒体受到过度的糖酵解的进一步保护。
- 因为任何没有进入TCA循环的丙酮酸都被导入柠檬酸盐,柠檬酸盐通过线粒体三羧酸载体分泌到胞浆中,并分解为乙酰辅酶A和草酰乙酸。草酰乙酸被转化为苹果酸,再被导入线粒体以维持回补,而乙酰辅酶a则是脂质生物合成和蛋白质乙酰化的前体。
- 尽管分解代谢TCA循环活性降低,但通过磷酸甘油脱氢酶活性和苹果酸转化为草酰乙酸的复合物I进入线粒体电子传递链的电子足以维持线粒体完整性和ATP生成。
-
减少形成的活性氧(ROS)从而对线粒体起到保护作用。
- 增殖细胞常常积累超过ATP合酶能力的电子传递流,从而形成过量的活性氧(ROS)。而过量的电子传递链产生ROS可能是癌基因诱导细胞衰老(OIS)现象的基础。OIS与氧化损伤和不可逆生长停滞相关。
- 丙酮酸脱氢酶(PDH)是一种将丙酮酸转化为乙酰辅酶A的限速酶,其活性的降低可降低OIS的产生并增强肿瘤的发生。
- 尽管过量的ROS对细胞生长和存活是有害的,但中等水平的ROS(在呼吸作用或以一种NADPH氧化酶的靶向方式生成)是一个重要的信号输入,有助于维持致瘤状态。
- ROS作为蛋白磷酸酶的抑制剂,如PTEN和PTP1B,以及Src家族激酶和MAPK的激活剂。
- 增加的ROS水平有助于激活HIF1A和NRF2转录因子,从而促进进一步促进肿瘤发生的转录程序。
癌细胞、增殖细胞中糖酵解的代谢中间产物
糖酵解在增殖细胞中用于生成许多生物合成过程的代谢中间产物。虽然糖酵解被描绘为介导丙酮酸生成的单一通路,但多个糖酵解中间体的可转移到其他通路,成为不同的生物合成的前体。
葡萄糖-6-磷酸
1. 通过磷酸戊糖途径,葡萄糖-6-磷酸部分氧化以生成NADPH和核糖-5-磷酸。
2. 该途径中的关键酶,转酮醇酶1(TKTL1)和转醛酶(TALDO)常在癌细胞中过表达。
3. 癌基因和抑癌基因都被证明调节PPP活性。
1. Ras诱导介导核糖5-磷酸生物合成的酶的转录上调。
2. 野生型p53通过直接结合和灭活PPP限速酶葡萄糖-6-磷酸脱氢酶(G6PD)来抑制PPP。果糖-6-磷酸
1. 氨基己糖途径将果糖-6-磷酸用作氨基己糖生物合成的底物
2. 氨基己糖生物合成的第一个关键步骤是谷氨酰胺果糖-6-磷酸氨基转移酶1(GFPT1)催化果糖-6-磷酸和谷氨酰胺生成葡糖胺-6-磷酸。
3. 通过产生N-乙酰葡糖胺(GlcNAc,N-和O-联糖基化的底物),氨基己糖途径为细胞糖基化反应提供底物(如硫酸乙酰肝素和透明质酸生物合成),增强受体介导的信号,调节选择蛋白(包括c-myc)的稳定性。磷酸二羟丙酮(DHAP)通过甘油醛-3-磷酸脱氢酶1(GPD1)为甘油-3-磷酸(用于合成不同的磷脂)。
-
3-磷酸甘油酸作为丝氨酸,甘氨酸生物合成的前体,并作为产生甲基供体基团和NADPH的手段。
丝氨酸的重要作用:
- 癌细胞可利用多达50%以上葡萄糖衍生的碳用于丝氨酸生物合成和其随后的代谢。限速丝氨酸生物合成的3-磷酸甘油酸脱氢酶(PHGDH),经常在乳腺癌和黑色素瘤中灶性扩增。
- 丝氨酸在细胞中具有独特的代谢作用,作为一碳单位(叶酸)的主要底物。为此,它的γ-碳可以被转移到载体分子四氢叶酸(THF)上,再通过线粒体中丝氨酸羟甲基转移酶2(SHMT2)和胞质溶胶中SHMT1催化反应,生成5,10-亚甲基-四氢叶酸和甘氨酸。然后5,10-亚甲基-四氢叶酸经历一系列的氧化还原转换,产生一碳-四氢叶酸。
- 一碳-四氢叶酸被用作嘌呤,胸苷,S-腺苷甲硫氨酸生物合成以及胞内甲基化反应的底物。此外,一碳-四氢叶酸产生多达50%的胞内NADPH。因此,缺氧驱动的SHMT2表达从缺氧相关的氧化应激中保护细胞。
- 亚甲基四氢叶酸脱氢酶2(MTHFD2,一碳途径的线粒体臂的组成部分)被认为是癌症中三个最经常过表达的代谢酶之一,说明一碳途径的改变普遍存在于肿瘤发生中。
肿瘤中糖酵解功能的平衡
为了平衡糖酵解的生物合成与其提供丙酮酸的作用,增殖细胞发展出调节丙酮酸激酶(PK,将磷酸烯醇丙酮酸转换为丙酮酸的调节酶)的机制。
肝肾之外(表达组织特异性PK亚型PKL和PKR),大多数组织表达PK的PKM的形式。 PKM存在两种剪接变异体。虽然PKM1生产丙酮酸更有效,但大多数增殖细胞和基本上所有癌细胞表达主要是PKM2变体。
PKM2的活动受到严格管制。PKM2通过酪氨酸磷酸化的抑制,由丝氨酸激活,PKM2也可以通过糖代谢的其它副产物的过量蓄积可变构激活,如SAICAR(核糖-5-磷酸生物合成的中间产物)。
以这样的方式,生长因子依赖性信号转导抑制PKM2,导致糖酵解中间体的累积,直到细胞生长需要的自由丝氨酸饱和的。通过实验引入该同种型PKM1影响丝氨酸依赖性生物合成,损害了体内癌细胞增殖和肿瘤发生,证实了PKM2的作用。
TCA循环中间产物用于生物合成
除了糖酵解中间产物外,生长因子信号还促进TCA循环中间产物用于生物合成前体的产生。
- PI3K/AKT的激活使细胞能够扩大乙酰辅酶A的胞浆池。为此,AKT激活ATP柠檬酸裂解酶(ACLY),该酶催化TCA循环衍生的柠檬酸盐在胞浆中局部裂解为乙酰辅酶A和草酰乙酸。
肿瘤中脂质合成代谢的作用:
- 正常成人组织中脂肪酸的从头生物合成水平较低,但肿瘤的发生与脂质生成的显著增加有关。从头合成脂质的能力的增加不仅有利于脂质双层的形成,而且还使细胞能够改变其膜组合物,以利于抗氧化损伤的饱和脂肪酸作为适应氧化应激的手段。
- 脂肪酸链的生物合成始于胞浆乙酰辅酶a经乙酰辅酶A羧化(ACC)生成丙二酰辅酶A,再经脂肪酸合成酶(FASN)组装成长脂肪酸链。
- 参与脂肪酰链生物合成的所有三个主要成分ACLY、ACC和FASN在转化细胞中经常上调,它们的抑制作用降低了体外和体内肿瘤细胞的生长。
- 除了脂肪酸的产生外,丙二酰辅酶A从头生物合成胆固醇也在肿瘤发生中起作用。通过抑制限速酶羟甲基戊二酰辅酶A还原酶(HMGCR)干扰胆固醇生物合成,恢复乳腺癌细胞的正常腺泡形态,表明膜成分和流动性的变化可能对组织结构和锚定生长产生影响。
- 除了提供柠檬酸盐外,TCA循环还为包括天冬氨酸和天冬酰胺在内的非必需氨基酸的生物合成提供代谢前体。天冬氨酸生物合成已被证明是严重依赖于细胞进行氧化磷酸化的能力。为了保持合成这些氨基酸的能力,它们的生产必须与回补底物的流入相平衡。
TCA循环回补底物:
1. 谷氨酰胺
1. 生长细胞的主要回补底物是谷氨酰胺。谷氨酰胺可以通过谷氨酰胺酶在线粒体中捕获,由此产生的谷氨酸通过谷氨酸脱氢酶或氨基酸转氨酶转化为α-酮戊二酸。大多数增殖细胞依赖谷氨酰胺的持续供应来维持TCA循环中间产物的完整性。
2. 尤其是在c-myc转化的细胞中,谷氨酰胺缺乏导致TCA循环的崩溃并触发细胞死亡,通过添加草酰乙酸或膜可渗透的a-酮戊二酸可以挽救细胞死亡。
3. 谷氨酰胺衍生的α-酮戊二酸氧化成草酰乙酸不仅有助于维持细胞合成柠檬酸盐的能力。
4. 同时,所产生的草酰乙酸可以转化为苹果酸,而苹果酸又可以通过苹果酸酶(ME1)进一步氧化成丙酮酸盐,其反应以独立于葡萄糖的方式产生NADPH。
5. 最后,α-酮戊二酸可在低氧或某些致癌导致葡萄糖衍生的乙酰辅酶A缺陷的背景下,为脂肪生成提供柠檬酸来源。在一个称为还原羧化的过程中,TCA循环的一部分会自我逆转,将α-酮戊二酸转化为柠檬酸盐。
2. 葡萄糖
1. 谷氨酰胺衍生的a-酮戊二酸不是唯一可以被增殖细胞利用的回补性底物。在人类患者和胶质母细胞瘤和非小细胞肺癌(NSCLC)小鼠模型中,TCA循环中间产物的主要来源不是谷氨酰胺,而是葡萄糖。
2. 在丙酮酸羧化酶(pc)的催化下,葡萄糖驱动的碳通过丙酮酸羧化进入tca循环,生成中间产物草酰乙酸。
3. 抗谷氨酰胺剥夺的胶质母细胞瘤细胞系衍生物在培养中的存活依赖于丙酮酸羧化酶。此外,即使存在大量谷氨酰胺,PC缺失的非小细胞肺癌细胞系抑制了脂肪酸生物合成,并显著减少增殖。说明细胞谱系背景,以及与细胞组织培养条件相关的次级效应,可能在回补底物的选择中发挥重要作用。
胞外乙酸盐用于生物合成
细胞外乙酸盐被一些癌症证明是乙酰辅酶A生物合成的来源。
- 各种局限于大脑的肿瘤类型,包括原发性胶质母细胞瘤以及起源于其他组织的脑转移瘤,已被证明吸收了外源性乙酸,并将乙酸衍生碳并入从头合成的脂肪酸中。
- 在乙酰辅酶A合成酶2(ACSS2,乳腺癌扩增的靶点)的催化下,胞外来源的乙酸盐转化为乙酰辅酶A。因此,乙酰基辅酶A的缺氧相关缺陷不仅可以通过α-酮戊二酸的还原羧基化得到增强,而且还可以通过提高血浆和间质液中游离乙酸的利用率来提高。
- 此外,ACSS2的上调也可能使细胞从组蛋白和其他细胞蛋白的脱乙酰化中释放出的乙酸循环。
4. 癌细胞增加对氮代谢的需求
与碳类似,氮代谢在肿瘤发生过程中经历复杂的重编程,生长信号提高了细胞对还原氮的需求。增殖细胞必须从头合成许多含氮分子,包括核苷酸、非必需氨基酸和多胺。
氮代谢的中间产物
-
谷氨酰胺是核苷酸生物合成中的关键结构构件(图5)。
- 谷氨酰胺水平被证明是细胞周期的限制因素,在某些细胞环境中,谷氨酰胺剥夺导致细胞周期在s期停滞。尽管谷氨酰胺在哺乳动物细胞中存在合成途径,但培养中的大多数增殖细胞需要外源性谷氨酰胺的供应。
- 一些细胞类型,如胚胎干细胞和管腔型乳腺癌细胞,在培养基中没有谷氨酰胺的情况下能够增殖,这表明在某些细胞环境中,谷氨酰胺可以从头合成。
- 植入免疫受损小鼠大脑的人类胶质母细胞瘤样本相对于正常脑组织积聚谷氨酰胺;此外,在c-met诱发肝癌的小鼠模型中观察到肿瘤组织中葡萄糖衍生谷氨酰胺的类似积聚。
- 谷氨酰胺合成酶(gs)在一些癌症中被发现过度表达。
-
精氨酸尽管是一种非必需氨基酸,但在某些致瘤环境中也成为必需的(图5)。
- 精氨酸是四个氮原子的载体,是多种含氮化合物的前体,其中包括多胺、肌酸、胍丁胺和脯氨酸生物合成的前体吡咯-5羧酸盐。
- 精氨酸的从头生物合成是尿素循环的一个组成部分,在精氨酸琥珀酸裂解酶(ASL)的催化下,精氨酸从精氨酸琥珀酸中衍生出来。精氨琥珀酸是由瓜氨酸和天冬氨酸的精氨琥珀酸合成酶(ASS1)产生的。
- ASS1和ASL酶在黑色素瘤、肾细胞癌和肝细胞癌中经常表观基因沉默。ASS1和ASL的表达减少与预后不良和化疗抵抗有关。
为什么肿瘤会放弃精氨酸的从头合成,转而依赖于其外源性内流?
- 一种可能是ASL和ASS1的失活使癌细胞积累鸟氨酸,然后用于生产多胺(一类含氮多阳离子脂肪族碳分子)。多胺已被证明能抑制凋亡并促进肿瘤生长和侵袭。
- 癌症中精氨酸营养不良的另一个可能原因可能与抑制ASS1驱动的精氨酸琥珀酸产生导致其底物天冬氨酸积累有关,天冬氨酸可用于核苷酸生物合成。
生长因子对氮代谢的调节
-
癌基因c-myc通过上调一些核苷酸生物合成酶的表达来调控核苷酸生物合成。
- c-myc调控的靶点包括催化嘌呤生物合成第一步的磷酸核糖焦磷酸合成酶2(prps2)和引发嘧啶环构建级联反应的氨甲酰磷酸合成酶II(cad)。
- 参与核苷酸生物合成的c-myc的其他靶点是胸苷酸合成酶(ts)和肌苷单磷酸脱氢酶1(impdh1)和2(impdh2)。因此,c-myc不仅促进了谷氨酰胺的吸收,而且促进了其在嘌呤和嘧啶碱生物合成中的应用。
- 谷氨酰胺酶在癌细胞中以依赖c-myc的方式频繁上调。
-
除了对谷氨酰胺代谢的影响外,c-myc还参与了非必需氨基酸代谢的重塑,包括脯氨酸代谢。
- 脯氨酸可以由谷氨酸或精氨酸衍生的鸟氨酸通过一种常见的中间体吡咯-5-羧酸盐产生。脯氨酸生物合成中主要酶吡咯-5-羧酸还原酶(PYCR1)的表达被c-myc上调。事实上,PYCR1是肿瘤中最常见的过度表达基因之一。
- 脯氨酸氧化酶(POX)介导脯氨酸降解,通过miR-23b由c-myc负控制,并且也是p53靶点。POX的表达已被证明通过在G2期触发细胞周期停滞来抑制肿瘤细胞的生长。
- 脯氨酸代谢改变对肿瘤发生的作用尚待阐明。有一种解释认为,升高的脯氨酸库可能促进胶原蛋白的产生和新的细胞外基质的沉积,促进肿瘤的侵袭。
除了c-myc的调控输入外,突变的p53等位基因也被证明有助于核苷酸生物合成基因的表达,其中包括impdh1和impdh2、GMP合成酶(gmps)、核苷挽救酶脱氧胞苷激酶(DCK)和胸腺嘧啶激酶(TK1)。
CAD酶的活性通过MAPK和MTORC1依赖性s6激酶的磷酸化调节。以这种方式,MTORC1驱动的CAD激活可使细胞响应细胞内谷氨酰胺的水平调节嘧啶的生物生成,因为谷氨酰胺有助于MTORC1的激活。
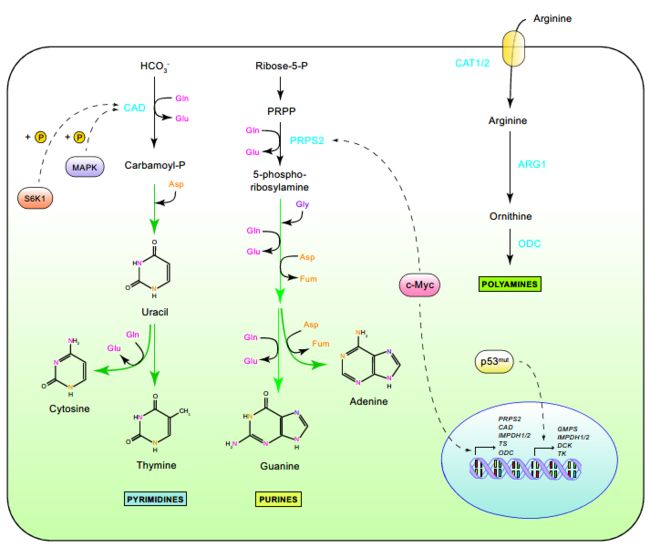
5. 代谢物驱动基因调控的改变
驱动肿瘤发生的异常激活的生长和存活信号有助于癌细胞代谢的重新编程,以促进营养的获取和生物合成。然而,代谢网络本身不仅是生长信号的被动接受者,相反,它直接将有关细胞代谢状态的信息传递给多种调节酶,其中包括那些介导染色质表观遗传标记沉积和去除的酶(图6)。
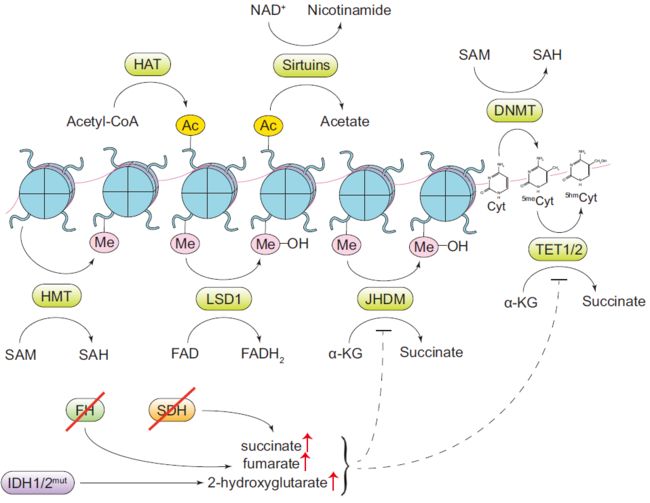
胞浆乙酰辅酶A对组蛋白乙酰化的调控
当细胞代谢的葡萄糖超过生物能量支持所需时,产生的一种关键代谢物是胞浆乙酰辅酶A。胞浆乙酰辅酶A是乙酰化组蛋白和其他蛋白质的酶的专一底物。
- 乙酰标记在组蛋白上的沉积与基因组DNA组装转录复合物的可接近性增加有关,并且具有快速的周转率,同时组蛋白乙酰化对细胞营养和信号状态的变化非常敏感。
- 葡萄糖的提取和再利用,以及通过引入致癌的KRAS或AKT突变体激活致癌信号,都会增加总组蛋白乙酰化,进而促进增强和更广泛的基因表达。
- AKT激活水平与整体组蛋白乙酰化状态密切相关。活化的akt通过激活acly,将胞浆柠檬酸盐转化为乙酰辅酶a和草酰乙酸,从而扩大了线粒体外的乙酰辅酶A。由葡萄糖和癌基因驱动的全局性组蛋白乙酰化增加都被acly抑制所阻断。
- p300组蛋白乙酰转移酶最近也被证明利用巴豆酰辅酶A作为底物。在组蛋白尾部的选定氨基酸残基上沉积巴豆酰标记比乙酰标记更能激活基因表达。
巴豆酰辅酶A可以由赖氨酸和色氨酸以及短链脂肪酸丁酸的分解代谢产生。
s-腺苷蛋氨酸对甲基化的调控
细胞中的许多甲基化反应,包括组蛋白尾部甲基标记的沉积、DNA上的胞嘧啶甲基化和mRNA上的腺苷甲基化利用s-腺苷蛋氨酸(SAM)作为甲基基团的供体。一些报告表明,组蛋白和DNA甲基化对SAM水平的改变敏感。
NAD+或FAD对清除乙酰和甲基标记的调控
清除乙酰和甲基标记受细胞代谢状态的指导。sirtuins,一类催化从组蛋白和非组蛋白中去除乙酰标记的脱乙酰酶,利用NAD+作为辅助因子, FAD作为赖氨酸特异性去甲基化酶lsd1的辅助因子。这些酶对NAD+或FAD有效性的变化敏感,它们协调了促进能源节约的全局性翻译后和表观遗传变化。
α-酮戊二酸对细胞翻译后修饰的调控
细胞中的多种翻译后修饰是由一大类α-酮戊二酸依赖的双加氧酶的成员进行的。α-酮戊二酸依赖的双加氧酶包括DNA脱甲基酶的TET家族,组蛋白去甲基化酶的Jumonji C家族,mRNA去甲基化酶FTO和ALKBH5,以及脯氨酰羟化酶(PHD)家族(响应于氧的可用性和氧化应激调节HIF1A水平)。
-
α-酮戊二酸依赖的双加氧酶的反应机制包括氧化α-酮戊二酸酯到琥珀酸盐中。因此,细胞内α-酮戊二酸水平可以直接影响双加氧酶的活性。此外,α-酮戊二酸依赖的双加氧酶易被其反应产物琥珀酸盐以及TCA循环中琥珀酸降解的下游产物富马酸盐抑制。
- 琥珀酸脱氢酶(SDH)的双等位丢失,一种在家族性副神经节瘤和嗜铬细胞瘤以及散发性胃肠道间质肿瘤的子集中发现的改变,会导致琥珀酸的积聚。
- 在家族性癌症综合征HLRCC(遗传性平滑肌瘤病和肾细胞癌)以及副神经节瘤和嗜铬细胞瘤中也发现富马酸代谢酶富马酸水合酶(FH)的丢失和富马酸盐的积聚。显示SDH和FH丢失的肿瘤具有许多与双加氧酶抑制一致的表型特征,其中包括DNA甲基化的特征性全球增加以及HIF1A水平升高。事实上,在sdh丢失的情况下,可以通过添加膜透性形式的a-酮戊二酸来逆转。
-
调节α-酮戊二酸依赖性双加氧酶活性的另一类与肿瘤相关的基因突变是异柠檬酸脱氢酶1(IDH1)和异柠檬酸脱氢酶2(IDH2)的功能增益突变。
- IDH1和IDH2的突变等位基因表现出一种不寻常的酶功能。与野生型异柠檬酸脱氢酶(将TCA循环代谢物异柠檬酸盐转化为α-酮戊二酸盐)不同,IDH的一种突变形式更喜欢使用α-酮戊二酸盐作为底物,催化其转化为2-羟基戊二酸盐(2-Hg)的D-对映体。由于其结构类似于α-酮戊二酸,2-Hg作为α-酮戊二酸依赖性双加氧酶的竞争性抑制剂。
- IDH驱动的胶质瘤、白血病和软骨肉瘤显示出显著的CpG岛高甲基化,这让人想起SDH和FH缺陷癌中的高甲基化表型。AML中的IDH突变与tet2-甲基胞嘧啶羟化酶的失活突变互斥,进一步暗示突变IDH是表观遗传重塑的真正驱动因素。
- 体内将突变的IDH1等位基因敲入正常造血细胞和软骨细胞会引发靶细胞系的异常扩张。在各种细胞环境中,突变idh等位基因的异位表达或外源性d-2-hg的治疗足以诱导dna和组蛋白甲基化并阻断细胞分化。
- 小分子介导的胶质瘤突变型IDH1和白血病突变型IDH2的抑制在这些情况下显著促进分化。据报道,idh1和idh2的小分子抑制剂可在治疗aml患者的Ⅰ期临床试验中诱导缓解。
- 一些肿瘤类型显示2-hg升高,即使没有idh突变。此外,发现LD2-Hg水平在缺氧反应中增加,牵涉到L- 2-Hg作为细胞应激反应缺氧的代谢调节剂。
6. 代谢与微环境的相互作用
细胞代谢状态的信息不仅影响其自身的长期决策,而且有可能影响其周围其他细胞的命运。许多遗传稳定的细胞类型,其中包括肿瘤相关的成纤维细胞、内皮细胞以及先天性和适应性免疫系统成分,由于居住在生长中的肿瘤附近而发生特征性表型变化。
肿瘤通常面临营养不良和缺氧的环境,并开发出各种营养清除策略来绕过这些限制。缺氧阻碍细胞进行氧化磷酸化和其他需要氧气的反应的能力,并破坏氧化还原平衡,影响细胞信号传导和转录程序。癌细胞与其微环境之间的相互相互作用会施加选择性压力,从而进一步影响癌细胞的代谢。
癌细胞如何对其微环境进行重新编程以帮助肿瘤生长和扩散?
这种重新编程包含多种策略,其中包括分泌生长因子、改变细胞外基质和细胞-细胞相互作用。除此之外,增殖的癌细胞也改变了周围细胞外环境的代谢组成(图7)。
-
肿瘤细胞对细胞外葡萄糖和谷氨酰胺的高利用导致了细胞外乳酸的积累,使其影响肿瘤微环境中的许多细胞类型。
- 乳酸水平的增加通过减弱树突状细胞和T细胞的激活和单核细胞的迁移促进免疫许可微环境的出现。
- 乳酸刺激常驻巨噬细胞极化至所谓的M2状态,在免疫抑制和伤口愈合中发挥作用。
- 乳酸的积累有助于促进血管生成。乳酸促进hif1a的稳定,激活内皮细胞中的nf-kb和pi-3激酶信号,并诱导肿瘤相关基质细胞分泌促血管生成因子vegf。
- 乳酸水平的增加也刺激成纤维细胞产生透明质酸,这可能有助于肿瘤的侵袭性。
- 乳酸通过单羧酸转运体mct1分泌到细胞外,与h+的共转运偶联,导致细胞微环境酸化。
-
线粒体脱羧反应中产生的过量二氧化碳也有助于细胞外酸化。
- 二氧化碳扩散到细胞外空间,在那里通过一类细胞外碳酸酐酶转化为H+和HCO3。酸酐酶,特别是CAIX异构体的表达,在缺氧期间升高。
- 增加的细胞外酸化刺激基质金属蛋白酶(mmps)和组织蛋白酶的蛋白水解活性,促进细胞外基质成分的降解并增强肿瘤的侵袭性。
-
一些肿瘤采用独特的策略促进其周围免疫许可微环境的出现。
- 许多固体肿瘤类型过度表达色氨酸二加氧酶吲哚胺-2,3-二加氧酶(IDO1)和色氨酸-2,3-二加氧酶(TDO2),它催化必需氨基酸色氨酸转化为其衍生物犬尿氨酸。随后色氨酸缺失会触发效应T细胞氨基酸剥夺相关的凋亡。此外,累积的犬尿氨酸作为芳香烃受体(AHR)的配体。
- 以依赖于AHR的方式,犬尿氨酸促进调节性t细胞表型,进一步促进抑制抗肿瘤免疫反应。最后,犬尿氨酸通过癌细胞自身的AHR增强自分泌信号,促进细胞外基质的降解和侵袭。
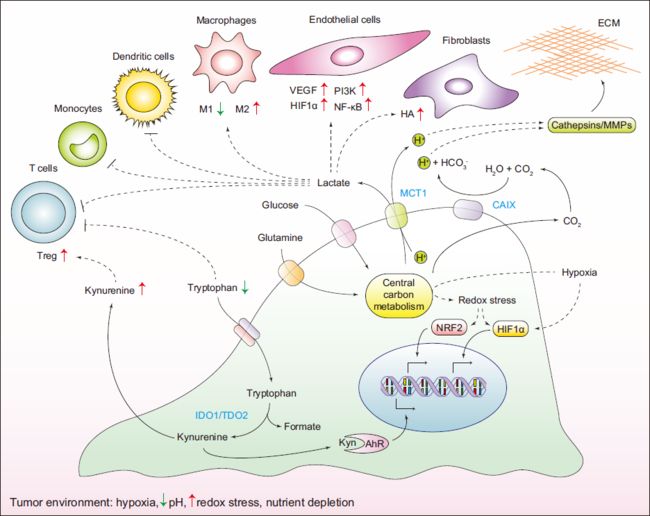
7. 癌细胞其他营养物质的代谢
目前为止的大多数研究都集中在葡萄糖和谷氨酰胺代谢的改变上,但癌细胞利用了许多其他营养物质,其中包括含硫氨基酸半胱氨酸和蛋氨酸、必需脂肪酸、胆碱、微量金属和维生素。
癌细胞中硫的代谢
多条证据表明硫的代谢可能在癌症中经历重新编程。
- 癌细胞通过xct转运体的转录上调上调来上调对半胱氨酸的摄取,并且继发于细胞对谷氨酰胺利用率的提高。
- 不少于30%的进口谷氨酰胺通过xct转运体以谷氨酸的形式离开细胞,同时被交换成半胱氨酸。
- 与癌细胞对半胱氨酸的大量利用相一致,半胱氨酸被发现是胰腺肿瘤中与正常胰腺组织相比消耗量第二大的氨基酸。
半胱氨酸在增殖细胞中有多种代谢产物,其中包括谷胱甘肽和铁硫簇的生物合成,以及硫化氢(H2S),其中的谷胱甘肽是氧化应激的保护剂,增加线粒体呼吸,防止细胞凋亡,促进血管生成。
肿瘤丝氨酸代谢
除了从头产生的丝氨酸外,来自外源性一碳供体的输入也可能影响一碳代谢和SAM水平。
- 饮食中缺乏丝氨酸和甘氨酸已被证明会损害小鼠结肠癌细胞的异种移植生长。
- 两种可以用作甲基供体的膳食化合物是胆碱及其衍生物甜菜碱。
- 在乳腺癌和前列腺癌中观察到胆碱摄取增加。饮食中摄入大量胆碱与致命前列腺癌风险显著增加有关。相反,高胆碱和甜菜碱饮食与乳腺癌和肺癌风险降低相关。此外,缺乏胆碱和甜菜碱的饮食被证明直接影响许多动物模型中的组蛋白和DNA甲基化。
- 外源性甲基供体的改变可能对细胞表观遗传状态有强烈的影响,从而有助于肿瘤的发生。
维生素和微量金属的代谢
微量营养素被用作各种酶的辅因子,在调节细胞代谢回路和信号转导方面发挥着复杂的作用。
- 抗坏血酸作为a-酮戊二酸依赖性双加氧酶的辅因子。抗坏血酸处理以依赖于TET2的方式影响胚胎干细胞和小鼠胚胎成纤维细胞的DNA甲基化。由于许多癌症的特征是CG岛甲基化增加,抗坏血酸输入的改变也可能因此影响其表观遗传状态。
- 与正常组织相比,癌细胞中选择的微量金属,如锌和铜的含量也发生了变化。尤其是在乳腺和卵巢肿瘤以及白血病中,铜的含量可以升高2到3倍。铜最近被证明是黑色素瘤细胞中由brafv600e驱动的erk激活所必需的,而铜耗竭的影响可以通过erk的结构性激活来挽救。
人体微生物群的代谢
最后,人体微生物群产生的广泛代谢物对肿瘤发生和发展的贡献才刚刚开始被阐明。
-
Pavlova, N. N. & Thompson, C. B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab 23, 27-47, doi:10.1016/j.cmet.2015.12.006 (2016). ↩
-
Birsoy, K., Wang, T., Chen, W.W., Freinkman, E., Abu-Remaileh, M., and Sabatini,D.M. (2015). An Essential Role of the Mitochondrial Electron Transport Chain in Cell Proliferation Is to Enable Aspartate Synthesis. Cell 162, 540–551. ↩